Производными эумеланина являются следующие цвета:
Black - черный
Blue - голубой
Seal - темно-коричневый, сил
Sable - соболиный
Chocolate - шоколадный
Chestnut - каштановый
Ruddy - дикий
Cinnamon - теплый светло-коричневый, коричный
Lilac - лиловый
Fawn - холодный бежевый, цвет олененка
Platinum - платиновый
O - Orange пигмент (среди любителей кошек обычно называется красным) - синтезирует желтую разновидность меланина - феомеланин. Диски пигмента О являются большими и более блестящими, чем диски пигмента В, что делает окрас очень ярким по всей длине волоса.
Если присутствует ген О, то ген В (или ген черного пигмента) инактивируется (т.е. остается неподвижным, неактивным). Из этого мы можем сделать вывод, что ген О доминирует над геном В. Эумеланин не формируется в фолликулах волос, оставляя возможность для формирования феомеланина. Красный, о котором мы сейчас говорим, является сцепленным с полом геном О, но не имеет никакого отношения к абиссинскому красному, который по существу не является красным вообще, а демонстрирует мутацию черного пигмента bl(корица).
Феомеланин формирует только 2 окраса: red & cream (красный и кремовый).
Если в хромосоме присутствует ген "о" (не красный) - в этом случае активируется ген В, - и в фолликулах волос формируются диски эумеланина. Ген "o" не формирует диски пигмента - он просто разрешает гену В стать активным. Это ген, который работает как переключатель: включить/выключить.
Ген "o" (не красный) записывается строчными буквами, но он не является рецессивным по отношению к гену О. Они являются одинаково доминантными.
РАЗВЕДЕНИЕ БРИТАНСКОЙ ПОРОДЫ
 
Вы занимаетесь разведением британской породы?
Если от двух короткошерстных родителей у вас рождается длинношерстный котенок, то вы задается справедливым вопросом: как это могло случиться?
Но вспомните приведенный выше пример 2, случай 1 и вы поймете как у вас мог родиться длинношерстный британский котенок.
Если в вашем британском питомнике родился длинношерстный котенок, вы можете быть абсолютно уверены, что оба родителя гетерозиготны по гену короткой шерсти (Ll) и это означает, что они оба в своем генотипе несут ген длинной шерсти (l). Если вы внимательно просмотрите родословные этих животных, вы наверняка найдете в предыдущих поколениях или перса, или представителя другой длинношерстной породы. Рецессивный ген может сохраняться в генотипе сколь угодно долго. Вероятность присутствия рецессивного гена в генотипе с каждым поколением будет уменьшаться, но теоретически это возможно на протяжении многих и многих поколений.
Поскольку появление длинношерстных котят не может нас радовать в британской породе, вам придется поменять производителя для вашей кошки.
Для введения гималайского окраса в британскую породу в основном использовались скрещивания с экзотами гималайского окраса. Последствием этого явилось введение рецессивного гена длинной шерсти (l) в британскую породу. И пройдет какое-то время, пока усилиями заводчиков влияние этого гена будет достаточно минимизировано. Хотя надо отдать должное - достойные представители британской породы пойнтового окраса - очень эффектны!
ГЕНЫ ПЛОТНОСТИ
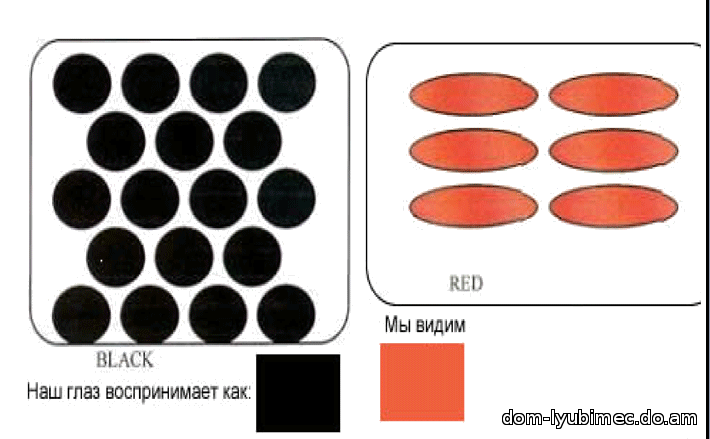
D - доминантный ген плотности. Гены пигмента, цвета и плотности работают совместно, чтобы создать окрас. При работе доминантного гена плотности D - диски плотно и ровно расположены вдоль длины волоса. Результатом работы гена D являются окрасы: black, seal, sable, chocolate, ruddy, cinnamon & red.
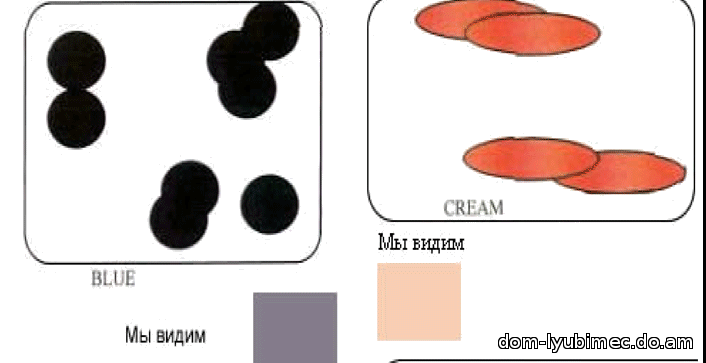
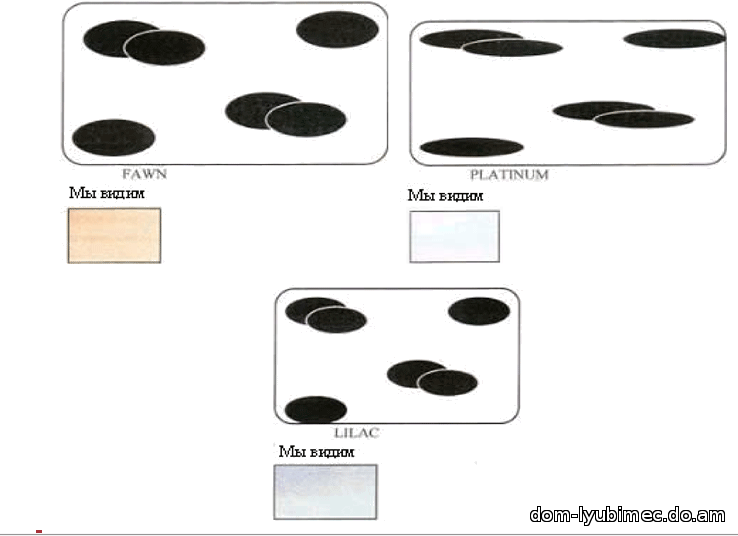
d - рецессивный или разбавленный ген плотности. Диски сгруппированы и разнесены. Результатом работы гена d являются окрасы: blue, lilac, fawn, platinum & cream.
На протяжении достаточно длительного времени из поколения в поколение разведенцев кочуют ошибки и заблуждения по генетике окрасов. Некоторые из них просто изумляют. Понимание этих заблуждений сделает вашу работу более понятной и интересной.
Одно из наиболее часто встречающихся заблуждений мнение, что голубой окрас является мутацией черного, или что вообще существует голубой или лиловый пигмент. Вспомните, все вы наверняка неоднократно слышали: "Мой кот является носителем голубого (или лилового) гена!"
Голубой не является мутацией черного и не изменяет форму диска, это все тот же неподвижный черный пигмент. Просто гранулы пигмента группируются, и расстояние между группами гранул увеличивается, а зрительно за счет более свободного прохождения и отражения света мы видим перед собой ослабленный, более светлый - голубой окрас. Голубой (так же как лиловый и светло-бежевый) являются результатом работы рецессивного гена "d ". Мутациями черного пигмента является ген b - chocolate и ген bl - cinnamon.
Приведенные в данной главе схемы-иллюстрации окрасов не совсем то, что мы в действительности видим под микроскопом, но они используются, чтобы наглядно продемонстрировать научную концепцию.
ГЕН СЕРЕБРА
I - ген ингибитор, который пресекает появление унаследованного окраса у прилежащей к коже части волоса. Этот ген отвечает за серебристую группу окрасов, за камео и за дымы.
i - рецессивный аллель гена-ингибитора, нормально проявляется окрас по всей длине волоса.
ГЕНЫ ЦВЕТА
C - доминантный ген полностью окрашенной шерсти, т.е. диски пигмента без изменения располагаются по всей длине волоса. Black, blue, chocolate, ruddy, cinnamon, fawn, seal, lilac, red, cream - примеры окрасов с полностью прокрашенной шерстью.
с - рецессивный ген цвета, создает различные формы диска, а также уменьшает количество дисков, расположенных на древке волоса. Бурманский, сиамский, тонкинский окрасы групп:
- Seal (соболиный, натуральная норка)
- Chocolate ( шампань, шампаньская норка)
- Blue (голубая норка)
- Lilac (платина, платиновая норка)
- примеры работы рецессивного гена "c". Все породы пойнтовых окрасов имеют рецессивный ген "c".
Следствием работы рецессивного гена "c" является и весь спектр альбиносов. Альбиносы появляются в результате дефицита пигмента, который выражается в отсутствии окраса кожи и шерсти, а, в зависимости от количества дефицита пигмента - в полном или частичном отсутствии пигмента в радужной оболочке глаз: розовые или голубые глаза и зрачок, который отсвечивает красным. Нормально окрашенные глаза отсвечивают зеленым.
Образование фермента тирозиназы определяется состоянием локуса "c". А тирозиназа является средством управления синтезом количества пигмента. Поэтому сиамское или бирманское высветление основного окраса появляется из-за того, что действительно происходит разбавление пигментов полностью окрашенной шерсти (ген С). Диски пигмента еще более удлиняются и отделяются друг от друга дальше, чем это можно было бы объяснить действием гена "d ".
В локусе С известно 4 рецессивных аллеля:
c - классический альбинос с розовыми глазами
cа - голубоглазый альбинос
Но в практической работе разведенцев интересуют следующие аллели гена "с":
сb - бурманский окрас, который изменяет форму дисков и расстояние между ними, что приводит к следующим изменениям в окрасах: черный превращается в соболиный, коричневый в шампань, а лиловый в платиновый.
сs - сиамский окрас, который является пойнтовым окрасом и превращает черный цвет в котиковый, а коричневый - в шоколадный.
Как ген сb , так и ген сs образовывают более бледные цвета вообще, чем их аналоги в доминантном гене цвета С. Хотя очень неожиданные результаты в разведении, связанные с затемнением шерсти, - могут дать полигены.
Итак, за окрас шерсти отвечает три типа генов: пигмента, цвета и плотности. При поверхностном рассмотрении кажется, что рецессивный ген "с" и рецессивный ген "d " выполняют одни и те же функции: увеличение промежутка между дисками пигмента. Однако необходимо понять и помнить: "с" и "d " два совершенно различных гена.
Мы в этой главе дважды вспоминали о шоколадном окрасе: однажды как "b" и однажды как "сs". Ген "сs" отвечает за пойнтовый окрас, однако мутация 'chocolate' одна и та же. Сплошной шоколадный окрас, шоколадный табби, затененный шоколад - все они являются представителями 'шоколадной' мутации гена "b" без гена пойнтового окраса "сs", они имеют доминантный ген полного цвета С.
Ген С оставляет ген плотности D или d без изменений. Диски пигмента ровно и аккуратно размещаются по всей длине волоса, только форма диска черного пигмента - в случае шоколадного или коричного окраса - изменяется на удлиненную, эллипсовидную. Удлиненная форма диска осветляет окрас, который мы зрительно воспринимаем. Тот же самый эффект происходит и с геном Orange (вспомните: пигментные диски красного окраса удлиненной формы), но внешне эти изменения не выглядят так драматически.
В ослабленных окрасах (присутствие гена d), в которых также присутствует рецессивный ген с, - удлиненные диски еще реже располагаются по длине волоса, что приводит к еще более бледным окрасам. Например: платиновый окрас состоит из шоколадного пигмента (ген b), разбавленной плотности (ген d) и гена бурманского окраса cbcb , который воздействует на пигментные диски, удлиняя и прореживая их (как сорняки при посеве). При взаимодействии генов b, d, и C мы имели бы лиловый окрас, но при генах b, d, и cbcb имеем более светлый, холодный вариант окраса - платиновый.
| 




