| Меню сайта |
|
 |
| Статистика |
Онлайн всего: 1 Гостей: 1 Пользователей: 0 |
|
|
Главная » 2018 » Апрель » 05
ВВЕДЕНИЕ
 
Сегодня мы все наблюдаем стремительное развитие фелинологии. Появляются новые породы, улучшается тип животных. Основной вклад в этот процесс вносят заводчики. Однако без специальных знаний заводчик довольно часто похож на средневекового алхимика, наугад смешивающего металлы, - в надежде получить золото. Что ж, иногда удается добиться некоторых результатов, но сколько сил и времени на это уходит!
Зачем же блуждать в темноте, искать то, что уже давно найдено? К сожалению, однако, многие книги по генетике кошек рассчитаны на узкий круг специалистов, без фундаментальных знаний в области генетики заводчику не посильны и способны убить даже самое ревностное желание в чем-либо разобраться.
Много источников информации использовалось при подготовке данного материала, однако целью моей работы было не напичкать заводчика энциклопедической информацией, а прежде всего объяснить закономерности наследования окрасов, используя основы общей генетики, необходимые для понимания.
Я надеюсь, что моя работа будет интересна не только заводчикам, но и тем, кто просто влюблен в кошек.
Наука генетика полна незнакомых для нас понятий: аллель, ген, хромосомы, гомозиготный, гетерозиготный, доминантный, рецессивный, полигены, генотип, фенотип.
Без понимания этих терминов невозможно использовать генетику в своей работе. Первоначально мы нуждаемся в знании общих законов, которые едины для всего живого.
Начнем с определения основных терминов генетики.
Хромосома - является системой линейно сцепленных генов, обеспечивающих хранение и передачу информации.
Ген - наименьший участок хромосомы, обусловливающий синтез определенного фермента. Основная единица наследственности для данного признака.
ДНК - дезоксирибонуклеиновая кислота. Хромосомы состоят из молекул ДНК и белка с примесью хромосомной РНК. Важным носителем наследственности является именно ДНК.
Аллели - альтернативные формы одного и того же гена, определяющие альтернативные признаки.
Локус - местоположение аллеля в хромосоме.
Гомозигота - несет согласованную пару генов для определенной характеристики, которая может быть как в доминантном, так и в рецессивном состоянии: AA или aa.
Гетерозигота - несет две различных характеристики в паре генов. Обычно один доминантный и один рецессивный ген: Аа.
Генотип - генетическая информация, заложенная в организме, комбинация всех аллелей индивидуума. Генотип как правило не соответствует внешнему проявлению.
Фенотип - есть внешнее выражение генотипа, но не выявляет весь генотип.
Доминантный - аллель, определяющий фенотип как в гомозиготном, так и в гетерозиготном состоянии.
Рецессивный - аллель, определяющий фенотип только в гомозиготном состоянии.
Другие термины будут объяснены в книге по мере необходимости.
К слову, количество пар хромосом у кота - 19, у человека - 23, у собаки -39!
Мы начнем с изображения: мы имеем 2 длинных потока генетического материала - в неделящихся клетках хромосомы представлены чрезвычайно длинными тонкими нитями, распространенными во всем объеме ядра:

Мейоз - это процесс деления клеточного ядра с образованием четырех дочерних ядер, каждое из которых содержит вдвое меньше хромосом, чем исходное ядро. Значение мейоза состоит в том, что он обеспечивает сохранение в ряду поколений постоянного числа хромосом у видов с половым размножением.
В процессе оплодотворения сливаются 2 половые клетки, каждая из которых несет одинарный набор. Следовательно, новообразованный организм получает одну хромосому от матери, а другую от отца.
Как гены передаются от родителей детям

Т.о. со сменой поколений происходит непрерывное перераспределение отцовских и материнских хромосом, - а значит, и заключенной в них генетической информации (окрас, телосложение, тип, здоровье, - одним словом - все, включая тот факт, что это котенок, а не лягушонок).
Некоторые гены являются доминантными (они проявляются в фенотипе), а некоторые являются рецессивными (они существуют, но не дают о себе знать в присутствии доминантного гена).
Если оба гена доминантные, то котенок проявляет эту характеристику в фенотипе и именно ее передает по наследству.
Если один ген доминантный, а другой рецессивный, то в фенотипе проявляется доминантная характеристика, а по наследству с вероятностью 50% передается как доминантная, так и рецессивная характеристика.
Если котенок имеет 2 рецессивных гена, то именно эту характеристику он проявляет в фенотипе и передает ее по наследству.
То, что мы видим во внешнем проявлении - называется фенотипом, а что кот несет генетически (как видимое, так и невидимое) - называется генотипом.
Запомните: генотип = ген
Мы познакомились с основными определениями, которые имеют отношение ко всей генетике. Но для заводчиков не менее интересен другой, более комплексный фактор, который также имеет большое значение в вашей разведенческой программе. Это - полигены.
Многие из самых заметных признаков организма представляют собой результат совместного действия многих различных генов: эти гены образуют особый генный комплекс, называемый полигенной системой. Хотя вклад каждого отдельного гена, входящего в такую систему, слишком мал, чтобы оказать сколько-нибудь значительное влияние на фенотип, почти бесконечное разнообразие, создаваемое совместным действием этих генов (полигенов), составляет генетическую основу непрерывной изменчивости.
Примером работы полигенов является рыжий окрас. Работа полигенов активно влияет на интенсивность, насыщенность окраса. Полигены, которые влияют на формирование рыжего окраса, - носят название RUFOUS или RUFUS (правомочны оба написания).
Однако, т.к. гены RUFOUS не являются частью гена ORANGE, - они в процессе деления могут быть наследованы любым цветным котом. Они могут проявиться обогащенным основным цветом на черном или голубом мраморе, равно как и на красном мраморе. Они дают яркость абиссинскому окрасу и насыщенность золотому. Они могут наследоваться через черного или голубого кота, проявившись в потомстве черных или голубых черепах.
Полигены являются наиболее таинственной и неизученной областью.
Полигены могут сказать нам практически все о конкретном коте, но мы, к сожалению, даже не имеем названия наибольшему множеству полигенов. Из-за того, что мы так мало знаем о работе полигенов, - мы зачастую можем пройти мимо необходимых нам знаний. Возможно многое, чего мы не понимаем в генетике кошек, - могло бы быть объяснено с помощью полигенов.

На диаграмме представлена зависимость рыжего окраса от количества и силы включенных в работу полигенов. Цифры на этой диаграмме являются произвольными.
Чем больше RUFOUS полигенов, тем насыщеннее красный окрас.
Просмотров:
368
|
Добавил:
k-9
|
Дата:
05.04.2018
|
ГЕНЕТИЧЕСКИЕ СИМВОЛЫ
 
Символы, используемые для генетического кодирования, очень удобны и позволяют вам легко стенографировать всю необходимую вам генетическую информацию.
Если подобно большинству людей вам легче начать с более простого - (а я делала именно так), мы начнем с генетических символов, которые определяют пол. По большому счету, пол и окрас - это основа генетики кошек.
Как у мужских, так и у женских особей во всех клетках имеются пары одинаковых (гомологичных) хромосом, но в одной паре хромосом они различаются. Это половые хромосомы. Эти хромосомы называют X- и Y- хромосомы.
XX - генотип кошки
XY - генотип кота
МЕНДЕЛЕВСКИЕ СХЕМЫ
Грегор Мендель родился в Моравии в 1822 г. В 1843 г. он поступил в монастырь августинцев в Брюнне (ныне Брно, Чехия), где принял духовный сан. Позже он отправился в Вену, где провел 2 года, изучая в университете естественную историю и математику, после чего в 1853 г. вернулся в монастырь.
Будучи в Вене, Мендель заинтересовался процессом гибридизации растений и, в частности, разными типами гибридных потомков и их статистическими соотношениями.
Краткое изложение сути гипотез Менделя:
- Каждый признак данного организма контролируется парой аллелей.
- Если организм содержит 2 различных аллеля для данного признака, то один из них (доминантный) может проявляться полностью, подавляя проявление другого (рецессивного).
- При мейозе каждая пара аллелей разделяется (расщепляется) и каждая гамета получает по одному из каждой пары аллелей (принцип расщепления).
- При образовании мужских и женских гамет в каждую из них может попасть любой аллель из одной пары вместе с любым другим из другой пары (принцип независимого распределения).
- Каждый аллель передается из поколения в поколение как дискретная неизменяющаяся единица.
- Каждый организм наследует по одному аллелю (для каждого признака) от каждой из родительских особей.
В разведении очень важно знать, что можно ожидать в потомстве от конкретных производителей. Для этого используются Менделевские схемы, которые позволяют определить, что теоретически мы можем ожидать в потомстве и в каких соотношениях.
Использование в разведенческой программе Менделевских схем является хорошим средством для прогнозирования и контроля вашей работы. Менделевские схемы не всегда могут ответить на все наши вопросы, но это происходит от недостатка статистической информации.
Обычно эти схемы называют "решеткой Пиннета" по имени биолога Punnett, который их изобрел для демонстрации принципов наследственности, разработанных Менделем. Но многие люди их называют непосредственно Менделевскими схемами.
Когда мы используем Менделевские схемы, - это не значит, что наши расчеты будут в точности соответствовать результатам практической работы. Но мы всегда будем точно знать, что мы можем получить от конкретных производителей и, что еще более важно, - чего получить не можем.
На примере половых хромосом X и Y мы проиллюстрируем работу Менделевских схем:

Генотип кота - XY и он может предоставить любому котенку как X-, так и Y- хромосому. Генотип кошки - XX , и любому котенку она может предоставить только X-хромосому.
Как вы видите, теоретически мы имеем шанс 50% получить в потомстве котиков и 50% - кошечек.
Когда мы имеем две одинаково доминантных характеристики - соотношение всегда будет 2:2 или 50/50.
В дальнейшем мы научимся строить более сложные схемы, которые позволят нам приподнять завесу над тайнами генетики. Используйте эту книгу, чтобы проложить маршрут дороги, по которой вы собираетесь идти, и пусть ваш путь будет полезным и интересным
ЧТО ТАКОЕ ОКРАС?
Простой вопрос, не правда ли? Окрас - это цвет шерсти конкретного кота, которого мы видим перед собой.
Не торопитесь! Все не так очевидно! Окрас - это всегда составляющая из 3-ех компонент:
с отдельным геном для каждого.
Я позволю себе удовольствие сделать небольшое отступление в область анатомии и химии. Эти знания будут необходимы нам для понимания существенных моментов в генетике окрасов, а также в некоторых сопутствующих моментах (таких, как глухота у белых котов).
На очень ранних этапах развития у зародыша в области нервной трубки осуществляется закладка будущих пигментных клеток. Сами они производить пигмент еще не могут, для этого они должны принять веретенообразную форму, пригодную для миграции. Клетки, синтезирующие пигмент, называются меланоцитами. Меланоциты мигрируют от нервной трубки к фолликулам волос, коже, нервной системе, среднему уху и глазам. Меланоциты синтезируют фермент тирозиназы, который через длинную цепь превращений производит два вида меланина: черный, который называется эумеланином и желтый - феомеланин.
Если вы не все поняли в предыдущем абзаце, - не беспокойтесь. Главное, что мы теперь знаем: есть некоторые химические превращения, которые оказывают влияние на формирование окраса.
Теперь, вернувшись с нашего урока химии, мы постараемся научиться правильно записывать генетические формулы. Еще раз повторим:
- "Пигмент" - это непосредственно химическое соединение, которое содержится в окрашенном диске.
- "Цвет" - это форма и способ упаковки пигментных дисков по длине волоса.
- "Плотность" - это сгруппированность или разряженность дисков по длине волоса
Чтобы понять генетическую формулу окраса, понять, какие сюрпризы скрываются под тем или иным окрасом - мы должны понять взаимодействие этих генов.
Окрас шерсти - это всегда взаимодействие трех генов:
пигмента, цвета и плотности!
В генетических формулах доминантные гены записываются заглавными буквами, а рецессивные гены - строчными. Вначале всегда записывается доминантный ген, а затем рецессивный. Гены в генетических формулах обычно записываются в алфавитном порядке. В большинстве случаев строчными буквами обозначаются мутантные гены (так коричневый пигмент b является мутацией черного пигмента B). Доминантные характеристики почти всегда ближе к "дикому", природному типу, а рецессивные характеристики являются мутациями, однако некоторые доминантные характеристики также являются мутациями.
Гены пигмента, цвета и плотности расположены в разных местах хромосомы, но всегда присутствуют в генотипе любого кота (даже белого).
Вначале рассмотрим гены пигмента и гены плотности. Вперед!
РАЗВЕДЕНЧЕСКАЯ ПРАКТИКА
Как мы можем использовать полученные теоретические знания в разведенческой практике?
Давайте рассмотрим случай, если вы занимаетесь разведением экзотов.
ПРИМЕР 1: В вашем питомнике вы используете скрещивания между экзотами и персами, но не знаете: гомозиготны или гетерозиготны по гену короткой шерсти ваши экзоты (Ll или LL, поэтому генотип запишем следующим образом L -).
Предположим, что вы хотите повязать экзотическую кошечку (L-) с персидским котом (ll).
Случай 1: Кошка гетерозиготна по гену короткой шерсти: Ll.

В нашем случае мы получим экзотов и персов в соотношении 1:1, но все экзоты будут гетерозиготны по короткой шерсти и являться носителями длинной шерсти.
Примечание. В разных международных фелинологических организациях существуют различные правила по поводу регистрации длинношерстных потомков от скрещивания экзотов и персов. Например, в CFA такие котята регистрируются как длинношерстные экзоты по классу AOV (Any Other Variety - любые другие разновидности) и не допускаются к участию в чемпионате. Причем совершенно не обязательно иметь экзотического кота в родителях, - даже если экзот будет и в более отдаленных генерациях родословной, - ваш длинношерстный котенок будет считаться длинношерстным экзотом и зарегистрирован по классу AOV.
В других фелинологических организациях, например в FIFe, длинношерстный котенок от аналогичного скрещивания регистрируется как персидский, наравне с остальными персами.
Случай 2 : Кошка гомозиготна по гену короткой шерсти: LL.

В этом случае в помете мы получим только гетерозиготных по короткой шерсти экзотов, которые все будут являться носителями длинношерстного гена l.
Аналогичным примером является работа с сиамами и балинезами или ориенталами и яванезами.
В некоторых фелинологических ассоциациях короткошерстные котята, полученные в результате скрещивания короткошерстных и длинношерстных животных (все тот же пример скрещивания перса с экзотом или сиама с балинезом) называются Variants (вариация или привычное для нас слэнговое название "вар"). Наверняка многие из вас в родословных животных встречали следующее обозначение окраса: EXO d Var. Это означает, что данный котенок получен от скрещивания длинношерстного и короткошерстного родителей - и ничего больше!
ПРИМЕР 2: В вашем питомнике вы хотите повязать между собой двух экзотов, но не знаете гомозиготны они по короткошерстному гену (LL) или гетерозиготны (Ll).
Случай 1: Кошка и кот гетерозиготны по гену короткой шерсти: Ll.

Теоретически в помете мы получим экзотов и персов в соотношении 3:1, но 50% котят будут являться носителями длинношерстного гена, 25% котят будут гомозиготны по гену короткой шерсти и 25% котят - будут гомозиготны по длинношерстному гену.
К сожалению, мы не сможем определить по фенотипу кто же из котят будет гомозиготным экзотом. Но так же в результате такого скрещивания мы получим и длинношерстного персидского котенка.
ЗАКЛЮЧЕНИЕ: От скрещивания двух экзотов, гетерозиготных по гену короткой шерсти, может родиться длинношерстный персидский котенок.
Случай 2 : При скрещивании двух экзотов:
Кошка гомозиготна по гену короткой шерсти: LL.
Кот гетерозиготен по гену короткой шерсти: Ll.
В данном случае совершенно не принципиально кто из родителей (кот или кошка) будут гомозиготны или гетерозиготны по гену короткой шерсти. Просто для построения менделевской схемы мы делаем предположение, что гомозиготна по гену короткой шерсти будет кошка.

Все котята в помете будут экзотами и 50% из них будут гомозиготны по гену короткой шерсти.
Задача каждого заводчика - перевести необходимые и благоприятные для работы гены в гомозиготное состояние.
Все рассмотренные выше примеры справедливы для работы с сиамами и балинезами или ориенталами и яванезами.
ГЕНЫ ПИГМЕНТА
B - черный пигмент, отвечает за синтез разновидности пигмента меланина - эумеланин. Черный диск пигмента является совершенно круглым, толстым и действительно черным, если гранулы пигмента рассмотреть под мощным микроскопом.
Следующие рецессивные гены также вырабатывают эумеланин, но не круглой, а эллипсовидной формы, что изменяет и внешне воспринимаемый окрас волоса:
b - коричневый пигмент (называемый шоколадным или каштановым). Коричневый диск пигмента удлиненной эллипсовидной формы с размытыми контурами.
bl - светло-коричневый пигмент (так называемый цвет корицы). Светло-коричневый диск более удлиненной и утонченной формы, благодаря свободному отражению света от дисков выглядит более бледным, искрящимся и блестящим.
Просмотров:
404
|
Добавил:
k-9
|
Дата:
05.04.2018
|
Производными эумеланина являются следующие цвета:
Black - черный
Blue - голубой
Seal - темно-коричневый, сил
Sable - соболиный
Chocolate - шоколадный
Chestnut - каштановый
Ruddy - дикий
Cinnamon - теплый светло-коричневый, коричный
Lilac - лиловый
Fawn - холодный бежевый, цвет олененка
Platinum - платиновый
O - Orange пигмент (среди любителей кошек обычно называется красным) - синтезирует желтую разновидность меланина - феомеланин. Диски пигмента О являются большими и более блестящими, чем диски пигмента В, что делает окрас очень ярким по всей длине волоса.
Если присутствует ген О, то ген В (или ген черного пигмента) инактивируется (т.е. остается неподвижным, неактивным). Из этого мы можем сделать вывод, что ген О доминирует над геном В. Эумеланин не формируется в фолликулах волос, оставляя возможность для формирования феомеланина. Красный, о котором мы сейчас говорим, является сцепленным с полом геном О, но не имеет никакого отношения к абиссинскому красному, который по существу не является красным вообще, а демонстрирует мутацию черного пигмента bl(корица).
Феомеланин формирует только 2 окраса: red & cream (красный и кремовый).
Если в хромосоме присутствует ген "о" (не красный) - в этом случае активируется ген В, - и в фолликулах волос формируются диски эумеланина. Ген "o" не формирует диски пигмента - он просто разрешает гену В стать активным. Это ген, который работает как переключатель: включить/выключить.
Ген "o" (не красный) записывается строчными буквами, но он не является рецессивным по отношению к гену О. Они являются одинаково доминантными.
РАЗВЕДЕНИЕ БРИТАНСКОЙ ПОРОДЫ
 
Вы занимаетесь разведением британской породы?
Если от двух короткошерстных родителей у вас рождается длинношерстный котенок, то вы задается справедливым вопросом: как это могло случиться?
Но вспомните приведенный выше пример 2, случай 1 и вы поймете как у вас мог родиться длинношерстный британский котенок.
Если в вашем британском питомнике родился длинношерстный котенок, вы можете быть абсолютно уверены, что оба родителя гетерозиготны по гену короткой шерсти (Ll) и это означает, что они оба в своем генотипе несут ген длинной шерсти (l). Если вы внимательно просмотрите родословные этих животных, вы наверняка найдете в предыдущих поколениях или перса, или представителя другой длинношерстной породы. Рецессивный ген может сохраняться в генотипе сколь угодно долго. Вероятность присутствия рецессивного гена в генотипе с каждым поколением будет уменьшаться, но теоретически это возможно на протяжении многих и многих поколений.
Поскольку появление длинношерстных котят не может нас радовать в британской породе, вам придется поменять производителя для вашей кошки.
Для введения гималайского окраса в британскую породу в основном использовались скрещивания с экзотами гималайского окраса. Последствием этого явилось введение рецессивного гена длинной шерсти (l) в британскую породу. И пройдет какое-то время, пока усилиями заводчиков влияние этого гена будет достаточно минимизировано. Хотя надо отдать должное - достойные представители британской породы пойнтового окраса - очень эффектны!
ГЕНЫ ПЛОТНОСТИ
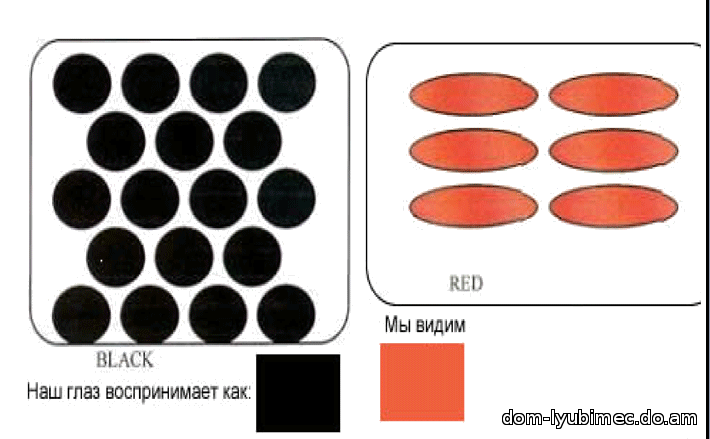
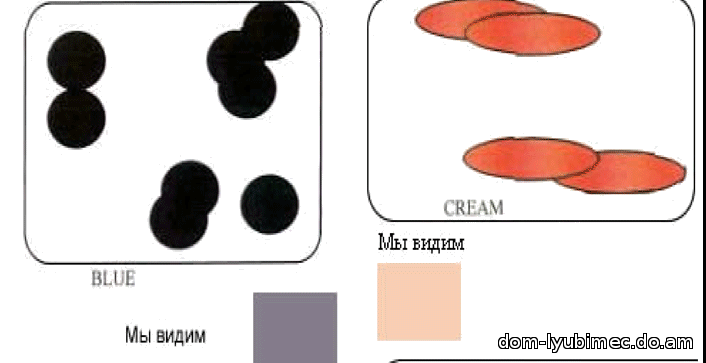
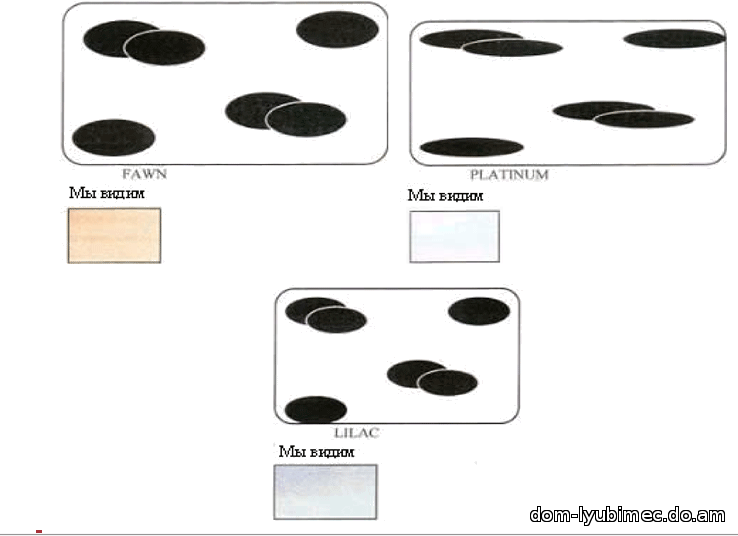
D - доминантный ген плотности. Гены пигмента, цвета и плотности работают совместно, чтобы создать окрас. При работе доминантного гена плотности D - диски плотно и ровно расположены вдоль длины волоса. Результатом работы гена D являются окрасы: black, seal, sable, chocolate, ruddy, cinnamon & red.
d - рецессивный или разбавленный ген плотности. Диски сгруппированы и разнесены. Результатом работы гена d являются окрасы: blue, lilac, fawn, platinum & cream.
На протяжении достаточно длительного времени из поколения в поколение разведенцев кочуют ошибки и заблуждения по генетике окрасов. Некоторые из них просто изумляют. Понимание этих заблуждений сделает вашу работу более понятной и интересной.
Одно из наиболее часто встречающихся заблуждений мнение, что голубой окрас является мутацией черного, или что вообще существует голубой или лиловый пигмент. Вспомните, все вы наверняка неоднократно слышали: "Мой кот является носителем голубого (или лилового) гена!"
Голубой не является мутацией черного и не изменяет форму диска, это все тот же неподвижный черный пигмент. Просто гранулы пигмента группируются, и расстояние между группами гранул увеличивается, а зрительно за счет более свободного прохождения и отражения света мы видим перед собой ослабленный, более светлый - голубой окрас. Голубой (так же как лиловый и светло-бежевый) являются результатом работы рецессивного гена "d ". Мутациями черного пигмента является ген b - chocolate и ген bl - cinnamon.
Приведенные в данной главе схемы-иллюстрации окрасов не совсем то, что мы в действительности видим под микроскопом, но они используются, чтобы наглядно продемонстрировать научную концепцию.
ГЕН СЕРЕБРА
I - ген ингибитор, который пресекает появление унаследованного окраса у прилежащей к коже части волоса. Этот ген отвечает за серебристую группу окрасов, за камео и за дымы.
i - рецессивный аллель гена-ингибитора, нормально проявляется окрас по всей длине волоса.
ГЕНЫ ЦВЕТА
C - доминантный ген полностью окрашенной шерсти, т.е. диски пигмента без изменения располагаются по всей длине волоса. Black, blue, chocolate, ruddy, cinnamon, fawn, seal, lilac, red, cream - примеры окрасов с полностью прокрашенной шерстью.
с - рецессивный ген цвета, создает различные формы диска, а также уменьшает количество дисков, расположенных на древке волоса. Бурманский, сиамский, тонкинский окрасы групп:
- Seal (соболиный, натуральная норка)
- Chocolate ( шампань, шампаньская норка)
- Blue (голубая норка)
- Lilac (платина, платиновая норка)
- примеры работы рецессивного гена "c". Все породы пойнтовых окрасов имеют рецессивный ген "c".
Следствием работы рецессивного гена "c" является и весь спектр альбиносов. Альбиносы появляются в результате дефицита пигмента, который выражается в отсутствии окраса кожи и шерсти, а, в зависимости от количества дефицита пигмента - в полном или частичном отсутствии пигмента в радужной оболочке глаз: розовые или голубые глаза и зрачок, который отсвечивает красным. Нормально окрашенные глаза отсвечивают зеленым.
Образование фермента тирозиназы определяется состоянием локуса "c". А тирозиназа является средством управления синтезом количества пигмента. Поэтому сиамское или бирманское высветление основного окраса появляется из-за того, что действительно происходит разбавление пигментов полностью окрашенной шерсти (ген С). Диски пигмента еще более удлиняются и отделяются друг от друга дальше, чем это можно было бы объяснить действием гена "d ".
В локусе С известно 4 рецессивных аллеля:
c - классический альбинос с розовыми глазами
cа - голубоглазый альбинос
Но в практической работе разведенцев интересуют следующие аллели гена "с":
сb - бурманский окрас, который изменяет форму дисков и расстояние между ними, что приводит к следующим изменениям в окрасах: черный превращается в соболиный, коричневый в шампань, а лиловый в платиновый.
сs - сиамский окрас, который является пойнтовым окрасом и превращает черный цвет в котиковый, а коричневый - в шоколадный.
Как ген сb , так и ген сs образовывают более бледные цвета вообще, чем их аналоги в доминантном гене цвета С. Хотя очень неожиданные результаты в разведении, связанные с затемнением шерсти, - могут дать полигены.
Итак, за окрас шерсти отвечает три типа генов: пигмента, цвета и плотности. При поверхностном рассмотрении кажется, что рецессивный ген "с" и рецессивный ген "d " выполняют одни и те же функции: увеличение промежутка между дисками пигмента. Однако необходимо понять и помнить: "с" и "d " два совершенно различных гена.
Мы в этой главе дважды вспоминали о шоколадном окрасе: однажды как "b" и однажды как "сs". Ген "сs" отвечает за пойнтовый окрас, однако мутация 'chocolate' одна и та же. Сплошной шоколадный окрас, шоколадный табби, затененный шоколад - все они являются представителями 'шоколадной' мутации гена "b" без гена пойнтового окраса "сs", они имеют доминантный ген полного цвета С.
Ген С оставляет ген плотности D или d без изменений. Диски пигмента ровно и аккуратно размещаются по всей длине волоса, только форма диска черного пигмента - в случае шоколадного или коричного окраса - изменяется на удлиненную, эллипсовидную. Удлиненная форма диска осветляет окрас, который мы зрительно воспринимаем. Тот же самый эффект происходит и с геном Orange (вспомните: пигментные диски красного окраса удлиненной формы), но внешне эти изменения не выглядят так драматически.
В ослабленных окрасах (присутствие гена d), в которых также присутствует рецессивный ген с, - удлиненные диски еще реже располагаются по длине волоса, что приводит к еще более бледным окрасам. Например: платиновый окрас состоит из шоколадного пигмента (ген b), разбавленной плотности (ген d) и гена бурманского окраса cbcb , который воздействует на пигментные диски, удлиняя и прореживая их (как сорняки при посеве). При взаимодействии генов b, d, и C мы имели бы лиловый окрас, но при генах b, d, и cbcb имеем более светлый, холодный вариант окраса - платиновый.
Просмотров:
344
|
Добавил:
k-9
|
Дата:
05.04.2018
|
ДРУГИЕ ГЕНЫ
 
Существует множество других символов, представляющих другие гены, но все они позволяют нам отождествить данного кота с его генотипом, который касается как его окраса, так и его физических характеристик. Когда вы смотрите на кота, - вы сразу же можете назвать состояние некоторых генов в его генетической формуле (например, - все доминантные гены), но ничего не знать о том, что данный кот маскирует или что несут его рецессивные гены (если вы не знаете его родителей или его потомков). Если мы знаем его родителей, - мы можем сделать приблизительную оценку относительно его генетической формулы. Но если мы знаем и его потомков - мы можем достаточно полно (в силу наших знаний на текущий момент) составить генетическую формулу нашего кота.
Для чего нам нужно знать генетические символы? Для того чтобы всегда уметь достаточно быстро расписать генетическую формулу наших котов, а с помощью Менделевских схем уметь прогнозировать ожидающие нас в нашем разведении результаты. Имея немного практики работы с генетическими формулами и Менделевскими схемами, - мы можем достаточно быстро стабилизировать те специфические характеристики, которые нам интересны.
Разумная генетика разрешает нам закреплять желательные в разведении характеристики и избегать неожиданных (чтобы не сказать отвратительных) характеристик. Перед тем, как составить план разведения - мы можем на листе бумаги вычислить, что нас ожидает в результате той или иной вязки. Это гораздо дешевле, чем покупать кота в надежде, что теперь-то мы сможем добиться того, о чем всегда мечтали.
Знание генетики может помочь нам использовать в разведении только перспективных для достижения наших целей котов. И тогда для осуществления наших разведенческих программ не нужно будет держать дома бесчисленное множество котов, ожидая каждый помет с надеждой, с какой мы каждый раз уповаем на джек-пот. А также мы получим огромное преимущество, снизив рождаемость племенной отбраковки. Также нам необходимы знания по генетике, чтобы избежать закрепления в генных пулах нежелательных (а иногда попросту опасных) рецессивных характеристик. Но это уже тема для специальной книги, а эта книга должна предоставить вам достаточные генетические знания, чтобы вы могли работать и со специальными книгами.
Генный пул - это сумма генотипов, обуславливающая проявление определенных фенотипов (как его качественных характеристик, так и количественных); это полный набор генов для данной породной группы. Сегодня, к сожалению, мы можем говорить только о генных пулах - более или менее изученных и доступных для разведенцев внутри избранных пород. Например, персы имеют достаточно большие генные пулы, а вот менее изученные породы, такие как Havana Brown, имеют гораздо менее доступные в разведении генные пулы. Как долго мы должны работать, чтобы избавиться в своей породе от нежелательных характеристик? Возможно ли разведение без мутаций? Есть бесчисленное множество различных мнений и статистических исследований на этот счет. Однако все они сводятся к тому факту, что человеческой жизни не хватит, чтобы решить все наши разведенческие проблемы и достоверно ответить на все наши разведенческие вопросы. Только знания, опыт, терпение и любовь могут помочь нашему продвижению вперед. И пусть иногда нам улыбается удача!
ГЕНЫ ТАББИ
А - Агути (перемежающиеся полосы основного и желтого цвета по всей длине волоса, действуют только на не оранжевый (non-orange) пигмент эумеланин). Агути является основой для всех окрасов табби. Его действие проявляется в широких разводах по основному окрасу, а так же, в меньшей степени, в полосах и пятнах.
а - нон-агути (отсутствие полос желтого цвета по длине волоса). Нон-агути не может действовать на ген О или Orange пигмент, поэтому все красные или кремовые кошки выглядят в большей или меньшей степени табби. "Все красные есть табби!" Это положение гораздо более подробно мы будем разбирать в Главе "Основы окраса шерсти". А сейчас мы сделаем небольшую попытку продолжить знакомство с некоторыми генетическими символами и понять, что они обозначают.
Следующие гены табби окрасов приводятся в порядке их доминантности.
Ta - тиккированный табби, иногда называемый абиссинским табби в честь породы абиссинских кошек, наиболее ярко демонстрирующих данный тип окраса, однако не имеющий ничего общего с породой как таковой. Ген, названный абиссинским, может быть представлен у любой породы, при любой текстуре и длине шерсти. Тиккинг выражен явно, четко, практически без рисунка, остаточные полосы (в зависимости от породы и стандарта) присутствуют на голове, шее, ногах и хвосте.
T - макрель (или тигровый) табби, вертикальные полосы на туловище в сочетании с общими для всех рисунчатых окрасов элементами: отметина "М" на лбу, ожерелья на груди, два ряда двойных пятнышек вдоль живота и полосы на хвосте и ногах.
tb - мраморный или классический образец табби, яркие полосы и разводы по корпусу, "бабочка" на плечах.
t - пятнистая или разорванная табби маркировка. Хорошо очерченные, круглые или овальные пятна на более приглушенном основном фоне. Многие разновидности диких кошек имеют специальный ген для пятнистого табби, но большинство исследователей считают, что домашняя кошка не имеет этого гена. Они считают, что пятна получаются из-за того, что тигровый рисунок разрывается в следствии влияния полигенов.
МЕНДЕЛЕВСКИЕ СХЕМЫ
Основной блок складывается из четырех квадратов и включает в себя 2 характеристики:
- кот - кошка (XX & XY)
- голубой - черный
- белый - сплошной
Чтобы проиллюстрировать пример работы с Менделевскими схемами, воспользуемся такими характеристиками, как голубой и черный. Пусть в нашем примере отец и мать будут оба черные, которые несут ген разбавленной плотности d . Генотип каждого BB/Dd. Двумя заглавными буквами BB обозначаем наличие черного пигмента - как в противоположном случае строчными буквами bb обозначали бы коричневый или шоколадный пигмент. Заглавной буквой D - обозначаем доминантный ген плотности и строчной d - рецессивный ген разбавленной плотности. В случае, когда оба родителя гомозиготны по одному и тому же гену (BB) - в построении Менделевских схем мы этот коэффициент можем не рассматривать, т.к. все потомство будет иметь тот же генотип и нам вовсе не обязательно проводить построение схемы, чтобы убедиться в этом. Чтобы сделать схемы наглядными и полезными в работе - всегда опускают любые генетические коды, которые не могут выдать переменный результат.
Каждый квадрат в нашей схеме имеет один генетический код от отца и один от матери. Схема выглядит следующим образом:

Теоретически в помете должно быть 3 черных котенка и 1 голубой. Это классическое Менделевское расщепление в отношении двух гетерозиготных форм. Это соотношение справедливо для любого множества признаков, которые контролируются одним доминантным и одним рецессивным геном. Мы прекрасно понимаем, что все вероятностные предсказания сбываются только при большом количестве испытаний и, взяв наугад 10 котят, мы среди них можем не обнаружить ни одного черного котенка. Но для нас важно другое: мы можем совершенно точно сказать, что и с какой вероятностью мы можем получить от данной вязки. Мы можем получить черного гомозиготного котенка, 2-ух черных гетерозиготных (носителей голубого) котят и голубого котенка в генетическом соотношении 1:2:1.
Это соотношение можно использовать и у людей для любого признака, который контролируется одиночным геном. Может ли человек скручивать язык трубочкой или нет? Имеют ли уши свободные доли или подсоединяются непосредственно к лицевой стороне? На эти вопросы точно также смогут дать ответы знакомые нам Менделевские схемы. Но вернемся все-таки к нашим котам.
Если вы вяжете гомозиготного черного кота с голубой кошкой (а она является гомозиготной по умолчанию!) - из всей схемы вам вообще понадобится только один квадрат:

Совершенно бесполезно рисовать много квадратов, если генотип всех котят будет Dd - черные, носители рецессивного гена разбавленной плотности.
Просмотров:
344
|
Добавил:
k-9
|
Дата:
05.04.2018
|
В разведенческой практике гомозиготное состояние генов называется "правильным". DD всегда произведет котенка основного окраса, dd всегда произведет котенка, который, по крайней мере, несет за собой ослабленный окрас. Dd в потомстве может передать как D, так и d. Кот с одним доминантным и одним рецессивным геном для данной характеристики называется гетерозиготным и не является примером "правильного" состояния генов. Кот будет внешне демонстрировать свою доминантную характеристику, но сможет передавать по наследству как доминантную характеристику, так и рецессивную. Если мы имеем черного кота носителя ослабленного окраса, то он будет фенотипически черным, но в генотипе иметь гетерозиготу Dd и передавать по наследству как D (основной окрас), так и d (ослабленный окрас).
С другой стороны, мы должны четко знать, что голубой кот может произвести только голубое потомство. Если в генотипе присутствует хотя бы один ген D, - кот будет черным. Истории, которые вы можете услышать, что от двух голубых родителей родился черный котенок, мягко говоря, являются некорректными. Наверняка или была вязка не с тем котом, или есть ошибка в родословной, или кто-то просто неправильно записал окрас родителя. Возможно, такая редчайшая мутация и произошла, но помните: коэффициент природной мутации - один на миллион!
Ситуация с красным и кремовым окрасами совершенно идентична. Давайте рассмотрим схему вязки двух красных гетерозиготных по гену плотности родителей. Кошка имеет генотип OO/Dd, а кот - OY/Dd (оба красные, носители гена разбавленной плотности). Красный окрас кота записываем OY , т.к. в его генетической формуле этому гену нет пары (более подробно смотрите раздел "Пол и ген O (Orange)").

Мы имеем знакомое нам соотношение 1:2:1. Один гомозиготный "правильный" красный котенок, два красных котенка носителя гена разбавленной плотности (крема) и один "правильный" кремовый котенок. По фенотипу соотношение 3:1.
А сейчас попробуйте решить небольшую генетическую задачу сами. Распишите, что мы получим в потомстве от вязки голубого кота (dd) и черной кошечки (Dd), которая является носителем голубого.

В литературе мы часто встречаем ссылки на гибриды F1 или F3 и т.д. Для генетического анализа наследования тех или иных признаков организма при половом размножении необходимо производить скрещивание двух разнополых особей. Скрещивание в генетике обозначают знаком умножения - x. При написании схемы скрещивания принято на первом месте ставить женский пол. Женский пол обозначают знаком (зеркало Венеры), мужской - (щит и копье Марса). Родительские организмы обозначают буквой P (первая буква латинского слова parenta - родители). Потомство от скрещивания двух особей с различной наследственностью называют гибридным. Гибридное поколение обозначают буквой F (первая буква латинского слова filii - дети) с цифровым индексом, соответствующим порядковому номеру гибридного поколения. Так, первое поколение будет F1, потомство гибридов первого поколения обозначают F2, третье поколение - F3 и т.д.
А вот теперь мы готовы познакомиться с новой информацией.
ГЕНЫ БЕЛОГО ФАКТОРА
 
W - ген обесцвечивания окраса, находится совершенно в другом месте хромосомы, чем гены B, C и D, и если ген W находится в доминантном состоянии, то гены окраса остаются в это время неподвижными. Ген W не является геном пигмента, этот ген называется геном белого фактора, который просто предотвращает разработку унаследованного цвета в древке волос. В дальнейшем мы часто будем использовать ссылку на "белый фактор".
w - не белый, нормально окрашенный. Фактически нет такого гена, который мы могли бы обозначить, как w, но символ w используется в генетическом кодировании, чтобы показать: есть или нет в генотипе W.
S - ген белой пятнистости или 'piebald' ген - пегий ген, который отвечает за формирование таких окрасов, как калико и би-колоры. Он может присутствовать в генотипе наряду с генами B, C, D и W.
s - полностью окрашенный (не би-колор).
G - ген отсутствия перчаток; "белые лапки" у такой породы, как священная бирма - вызваны действием рецессивного гена g.
g - ген наличия перчаток. Самостоятельный, независимый ген от гена S, отвечает за белые перчатки и шпоры. Большинство исследователей считают, что этот ген с давних времен получен в наследство от котов с сиамским окрасом и каким-то образом связан с геном cs, во всяком случае ген g встречается только либо в породах с пойнтовым окрасом, либо в породах, которые использовали котов с пойнтовым окрасом, как основу для работы или ауткросса.
ПОЛ И ГЕН O (ORANGE)
Помните, в начале нашей книги мы говорили, что пол и окрас - это основа генетики кошек. Это справедливо потому, что кошки являются совершенно уникальными созданиями в мире, (Мы, правда, об этом всегда знали и без генетики!), потому что у них окрас сцеплен с полом. Пол определяется X и Y хромосомами. Вы, конечно, помните, что генотип кошки - XX, а генотип кота - XY. Чтобы понять, как окрас сцеплен с полом, - нам придется рассмотреть реальную, фактическую форму хромосом X и Y. Даже по внешнему виду хромосомы напоминают написание самих букв X и Y. Y-хромосома по сравнению с X-хромосомой очень мала и большая часть локусов, характерных для Х-хромосомы, на ней отсутствует. Поэтому у особей мужского пола признаки, определяемые генами этого участка, проявляются по их состоянию на X-хромосоме, даже если они рецессивны. Это явление известно под названием наследования, сцепленного с полом. (Например, гемофилия, раннее облысение у человека).
У кошек ген O (Orange) расположен как раз на том участке X-хромосомы, у которого нет аналога на Y-хромосоме, и поэтому коты не имеют локуса для гена O (Orange) на Y-хромосоме. Котенок-кот имеет ген O или o на X-хромосоме, которую он унаследовал от матери. Если он имеет ген O, - он будет красным или кремовым, если он имеет ген o - он будет какого-то иного (не красного) окраса. И в разведении он передаст X-хромосому своим дочерям и Y-хромосому - сыновьям, определив тем самым пол котенка. Он не сможет передать ген O сыновьям, т.к. на Y-хромосоме в принципе нет локуса для гена О. Однако он прекрасно передаст и ген цвета С, и ген плотности D, и ген пигмента B как сыновьям, так и дочерям. Котенок-кошка, с другой стороны, получит по X-хромосоме от каждого из родителей. Обе X-хромосомы могут иметь О или о ген. Если она получит от родителей ОО, - она будет красного или кремового окраса, если она получит от родителей оо - ее окрас сформируется под действием пигмента В, но если она получит О от отца и о от матери (или наоборот), то она будет черепахового окраса, потому что О предоставит красный пигмент, а о позволит стать активным пигменту В. Таким образом ее окрас будет результатом смешения красного и черного цветов. Редкие представители котов черепахового или калико окрасов, как правило, имеют аномальное количество половых хромосом, (обычно XXY), и практически всегда стерильны.
Построим схему вязки красного кота и черной кошки. Генетическую формулу красного кота запишем следующим образом: O_/XY (пробелом обозначаем отсутствие гена Orange на Y-хромосоме). Генетическую формулу черной кошки запишем как oo/XX (по умолчанию предполагаем, что родители имеют в генотипе доминантный ген цвета СС, доминантный ген плотности DD и доминантный ген черного пигмента BB), т.к. сейчас хотим узнать, как распределятся по полу окрасы котят.
Косую черту "/" в генетических формулах я использую, чтобы разделить друг от друга различные характеристики. Это исключительно удобное изобретение в написании генетических формул, предложенное Дианой Браун. Мне кажется, что такой способ помогает более легкому прочтению и пониманию генетических формул.

Мы получаем котят-кошечек только черепахового окраса и только черного окраса котят-котиков. При таком разведении мы всегда получим только черепаховых кошек и только черных котов. Давайте поменяем окрасы родителей. Что же мы получим в потомстве? Только черепаховых кошек и только красных котов.
Все котята-кошки от одного красного и одного черного родителя будут черепахового окраса, но красная мать родит только красных сынов, а черная мать - только черных сынов. Теперь вы знаете, почему генетики говорят, что котенок-кот всегда наследует свой окрас от матери.
Если вы хотите строить ваши собственные схемы - вы должны четко знать, какие характеристики вам действительно интересны и необходимы. После этого вы должны записать генетические формулы двух интересующих вас котов. И как только вы имеете полностью записанные формулы, - вы берете по одному символу (гамете) из каждой пары и объединяете их во всевозможных комбинациях.
Например: Гетерозиготная белая кошка (Ww), маскирующая гетерозиготный по гену плотности черный окрас (Dd) - вяжется с точно таким же котом. Генетическую формулу записываем следующим образом: Ww/Dd. Существуют 4 возможные комбинации: WD, Wd, wD, wd. Если мы вяжем двух кошек с генотипом Ww/Dd, то схема будет выглядеть следующим образом:

А теперь декодируем (расшифруем) нашу схему:
- гомозиготный белый, маскирующий черный
- гомозиготный белый, маскирующий черный, носитель голубого
- гетерозиготный белый, маскирующий черный
- гетерозиготный белый, маскирующий черный, носитель голубого
- гомозиготный белый, маскирующий черный, носитель голубого
- гомозиготный белый, маскирующий голубой
- гетерозиготный белый, маскирующий черный, носитель голубого
- гетерозиготный белый, маскирующий голубой
- гетерозиготный белый, маскирующий черный
- гетерозиготный белый, маскирующий черный, носитель голубого
- гомозиготный черный
- гетерозиготный черный, носитель голубого
- гетерозиготный белый, маскирующий черный, носитель голубого
- гетерозиготный белый, маскирующий голубой
- гетерозиготный черный, носитель голубого
- гомозиготный голубой
Как вы видите, чтобы ответить на наши вопросы, - нам понадобилась схема из 16 квадратов. В потомстве теоретически мы можем получить 12 белых, 3 черных и 1 голубого котенка или, учитывая 2 характеристики: белое и черное, получим соотношение 12:4 или классическое 3:1. Среди окрашенных котов (2 характеристики: черное и голубое) мы снова имеем соотношение 3:1 - 3 черных и 1 голубой котенок.
Следующую схему изучите и попытайтесь декодировать самостоятельно, перед тем, как вы прочитаете список декодирования.
Задача: Рассмотрим пример скрещивания черной кошки CPC - колороносительницы (Ccs), гетерозиготной по гену плотности (Dd) с точно таким же котом. Генетическую формулу записываем следующим образом: Ccs/Dd. Существуют 4 возможные комбинации: СD, Cd, csD, csd. Если мы вяжем двух кошек с генотипом Ccs /Dd, то схема будет выглядеть следующим образом:

А теперь проверьте, так ли вы декодировали нашу схему:
- гомозиготный по гену полностью окрашенной шерсти, черный
- гомозиготный по гену полностью окрашенной шерсти, черный, носитель голубого
- колороноситель, черный
- колороноситель, черный, носитель голубого
- гомозиготный по гену полностью окрашенной шерсти, черный, носитель голубого
- гомозиготный по гену полностью окрашенной шерсти, голубой
- колороноситель, черный, носитель голубого
- колороноситель, голубой
- колороноситель, черный
- колороноситель, черный, носитель голубого
- колорпойнт, сил-пойнт
- колорпойнт, сил-пойнт, носитель голубого
- гомозиготный по гену полностью окрашенной шерсти, черный, носитель голубого
- гомозиготный по гену полностью окрашенной шерсти, голубой
- колорпойнт, сил-пойнт, носитель голубого
- колорпойнт, гомозиготный блю-пойнт
В потомстве теоретически мы можем получить 9 черных, 3 голубых, 3 сил-пойнта и 1 блю-пойнта котенка - известное нам соотношение 9:3:3:1. Или, учитывая 2 характеристики: сплошной и пойнтовый окрасы, получим соотношение 12:4 или классическое 3:1. Среди полностью окрашенных котов (2 характеристики: черное и голубое) мы снова имеем соотношение 3:1 - 9 черных и 3 голубых котенка и среди пойнтовых окрашенных котов (2 характеристики: черное и голубое) мы снова имеем соотношение 3:1 - 3 сил-пойнта и 1 блю-пойнт котенок.
ГЕНЫ ДЛИНЫ ШЕРСТИ
L - короткая шерсть, записывается как L, потому что короткая шерсть доминирует над длинной.
После недолгих размышлений мы приходим к забавному выводу: доминантный ген L поддерживает "длину" не "долго", и короткая "длина" доминирует над длинной "длиной"! (Этот факт многих обескураживает, когда они впервые знакомятся с генетикой, но нас с вами, уже знакомых с некоторыми генетическими превратностями - это не может сбить с толку и позволить запутаться в некоторых нелогичных на наш взгляд генетических закономерностях). Итак: L - доминантный ген короткой шерсти.
l - рецессивный ген длинной шерсти.
Просмотров:
489
|
Добавил:
k-9
|
Дата:
05.04.2018
|
ДРУГИЕ МУТАЦИИ
 
Известный английский биолог и генетик Рой Робинсон в одной из своих работ говорит: "Когда-то давно..., одной из первых мутаций, проявившихся у кошки - было появление черного окраса." Таким образом, все существующее ныне разнообразие окрасов является по большому счету результатом закрепленных природных мутаций знакомого всем дикого окраса агути. Но мутации касаются не только окраса или длины шерсти. Давайте сейчас немного поговорим о мутациях как таковых. Что это такое? Каким образом мутации проявляются? Что известно о мутациях у кошек?
Как мы знаем, генетика изучает два свойства органических форм - наследственность и изменчивость. Изменчивость является свойством, противоположным наследственности; она заключается в изменении наследственных задатков - генов. Существуют различные типы изменчивости. Изменение свойств и признаков организма может быть обусловлено изменением одного или нескольких генов под влиянием определенных условий среды. Такие изменения называются мутациями. Мутации возникают скачкообразно как новые качественные изменения. Например, вместо окраса шерсти агути - появляется белый окрас, вместо обычного уха - загнутое, и т.д.
Термин "мутация" впервые был предложен Г. де Фризом в его классическом труде "Мутационная теория" (1901-1903 г.г.). Мутацией он назвал явление скачкообразного, прерывного изменения наследственного признака. Основные положения своей теории Г. де Фриз в начале века сформулировал настолько точно и четко, что мне хотелось бы привести их полностью (тем более, что они до сих пор не утратили своего значения):
- Мутация возникает внезапно.
- Новые формы устойчивы.
- Мутации являются качественными изменениями.
- Мутации могут быть как полезными, так и вредными.
- Мутации могут возникать повторно.
В основе всех мутаций лежат биохимические процессы. Часто мутации называют по той функции, которую изменяет мутировавший ген (например, мутация, приводящая к торможению роста всего организма, так и называется - Dwarf (символ dw), карлик). Существуют мутации, нарушающие развитие нервной системы, повреждающие развитие конечностей, изменяющие развитие ушей или хвоста, образующие новые разновидности окрасов (или маскирующие существующие окрасы - как уже знакомая нам доминантная мутация, нарушающая способность пигментных клеток к миграции, Dominant White (символ W)), а так же влияющие на наличие, длину, курчавость и качество шерсти.
Давайте рассмотрим краткий список некоторых породообразующих мутаций:
M
отсутствие хвостаmнормальный хвост
Fd
сложенное ухоfdнормальные уши
Cr
закрученное ухоcrнормальные уши
HR
нормальная шерстьhrсфинкс (бесшерстность)
R
нормальная шерстьrкорниш рекс
Re
нормальная шерстьreдевон рекс
Wh
проволочная шерстьwhпроволочная шерсть
Rs
селкирк рексrsнормальная шерсть
В приведенном выше списке вы можете встретить несколько генетических кодов для обозначения нормальных ушей или нормальной шерсти. Такое обозначение распространено для удобной и понятной записи состояния аллелей. Когда мы говорим о какой-либо частной мутации шерсти, ушной раковины или хвоста - мы нуждаемся в символе, с помощью которого мы могли бы записать другую характеристику, или аллель, которая обязана присутствовать в парном наборе генов. Если кот гетерозиготный по данной мутации, должен быть символ, с помощью которого мы могли бы записать нормальную характеристику. Если кот гомозиготный, мы так же должны уметь это записать с помощью генетического кодирования, чтобы понимать с чем мы имеем дело.
Коты не имеют такого разнообразия в размерах, как собаки. Но они имеют многие мутации, которые встречаются у других домашних животных: кроликов, мышей, гвинейских свинок, собак.
Например, пойнтовый окрас шерсти встречается у гвинейских свинок, тушканчиков, хомяков, мышей, кроликов, крыс. (И у лошадей, между прочим, но это совершенно другая генетическая история...) Поэтому в принципе заводчики котов вполне могли бы ожидать проявления и других разновидностей окраса, свойственных вышеназванным животным.
Иногда мутация в качестве шерсти или окрасе может на первый взгляд показаться очень незначительной. Но заводчики всегда должны обращать внимание на необычных котят, если такие появляются в помете. И если что-то покажется вам действительно неординарным и новым, - вы должны попытаться закрепить и развить эту мутацию. Ведь с помощью селекции любая новая характеристика может быть закреплена и увеличена, а теоретически вы к этому уже готовы!
ИТОГИ
На этом мы пока завершим наш генетический ликбез. Впереди у нас с вами еще много интересного: мы познакомимся с формированием цвета глаз с точки зрения генетиков, рассмотрим основные типы (сплошные, табби, дымы, затененные, партиколоры, биколоры, пойнтовые) окрасов шерсти, подробно разберем различные окрасы в большинстве признанных пород, попробуем разобраться с генетически возможными окрасами. А пока попробуем все, с чем мы уже познакомились, собрать для наглядности в одной таблице:
Обозначение
НазваниеДоминантностьВозможные комбинацииФенотип
A
AgoutiДоминантныйAA AaОкрас шерсти с агути (с табби) образцом рисунком
a
Non-agoutiРецессивныйaaОкрас шерсти без агути (без табби) образца рисунка
B
BlackДоминантныйBB Bb BblОкрас шерсти чисто-черный. У гималаев, сиамов, бурм этот окрас называется Seal. У ориенталов этот окрас называется Ebony.
b
ChocolateРецессивныйbb bblОкрас шерсти шоколадный. У ориенталов этот окрас называется Havana.
bl
CinnamonРецессивныйblblШерсть полностью окрашена, без пойнтов.
C
Full ColorДоминантныйCC Ccs Ccb>Шерсть полностью окрашена, без пойнтов.
cb
Burmese pointsРецессивныйcbcb cbcsБурманское разбавление - окрас шерсти темно-коричневый (называется Sable), на более светлых окрасах пойнты заметны лучше. Пойнты имеют цвет основного окраса, но чуть темнее. Тонкинезский окрас.
cs
Siamese pointsРецессивныйcscsПойнтовый окрас шерсти. Маска на лице, ногах, хвосте, более светлое в тон тело, глаза ярко-голубые.
ca
Blue eyed AlbinoРецессивныйcacaБелый окрас шерсти со светло-голубыми глазами. Не путать с белым голубоглазым (ген W).
c
AlbinoРецессивныйccБелый окрас шерсти, цвет глаз без пигментации (просвечивает розовым).
Cu
Curled EarsДоминантныйCuCu CucuУши искривлены, завернуты к затылочной части. Встречается в породе Американский Керл.
cu
Straight earsРецессивныйcucuПрямые, нормальные уши.
D
Density, non-dilutedДоминантныйDD DdНе ослабленные окрасы шерсти: красный, черный, шоколадный, коричный.
d
DilutedРецессивныйddОслабленные окрасы: кремовый, голубой, лиловый, бежевый.
Fd
Folded earsДоминантныйFdFd FdfdСложенные складкой уши. Встречается в породе скоттиш-фолд.
fd
Straight earsРецессивныйfdfdПрямые, нормальные уши.
HR
Normal coatДоминантныйHrhr HrhrКот с обычной, нормальной шерстью.
hr
HairlessРецессивныйhrhrПрактически без шерсти, встречается у сфинксов.
I
InhibitorДоминантныйII IiПодавляет развитие пигментации в волосе, частично окрашено только окончание волоса, шерстинки с серебристо-белым основанием.
i
Non SilverРецессивныйiiОкрас шерсти без серебра.
L
ShorthairДоминантныйLL LlКороткая шерсть.
l
LonghairРецессивныйllДлинная шерсть.
А чтобы чувствовать себя спокойнее и увереннее при знакомстве с новым материалом - давайте еще раз повторим основы основ генетики окрасов.
Существует всего ДВА вида пигментов:
НЕТ генов пигмента для:
- BLUE
- CREAM
- LILAC
- FAWN
- PLATINUM
Существует ДВЕ мутации пигмента BLACK:
При изменении (мутации) гена сплошного окраса Сolor, но при сохранении пигмента Black, мы получаем:
- cb - SABLE
- cs - SEAL POINT
- cbcs - NATURAL (Tonkinese)
В результате работы гена разбавления d мы получаем:
- BLUE (когда пигмент BLACK)
- CREAM (когда пигмент RED)
- LILAC (когда пигмент CHOCOLATE)
- FAWN (когда пигмент CINNAMON)
- PLATINUM (когда пигмент SABLE)
Нет одного образующего окрас гена. Нет такого гена, который бы отвечал за образование голубого окраса. Точно так же, как и нет единственного (отдельного) гена, который бы отвечал за образование черного, или любого другого окраса.
Ослабленный окрас (Dilute) автоматически отнюдь не означает голубой окрас, как и доминантный - это вовсе не обязательно черный.
Если вам больше ничего не понадобится в этой книге, пожалуйста, запомните: необходимо ТРИ гена, чтобы получить ЛЮБОЙ окрас. Вы можете повторять как молитву: "Окрас - это пигмент, цвет и плотность! Аминь".
Диана Браун
авторизированный перевод Ольги Ракитных
http://rolandus.org
Просмотров:
338
|
Добавил:
k-9
|
Дата:
05.04.2018
|
ГЕНЕТИКА ОКРАСКИ У КРОЛИКОВ.
Источник: В.Г. Плотников, Н.М. Фирсова. Разведение, кормление и содержание кроликов. М. 1989 г.
Генетическое улучшение животных трудоемкий процесс. Для работы в этом направлении нужна наблюдательность и знание законов генетики. Потомки не бывают такими же как и их родители. Одни признаки более устойчиво передаются по наследству, другие требуют кропотливого и не всегда успешного подбора и отбора. Наследственная информация родителей содержится в половых клетках. Каждая половая клетка (яйцеклетка, сперматозоид) несет в себе одинаковое количество хромосом, в которых расположены гены- единицы наследственности, ответственные за те или иные признаки.
В половых клетках кролика 22 хромосомы (гаплоидный набор), а в соматических клетках 44 (22пары). Хромосомы содержат огромное количество генов, каждый из которых занимает определенный участок (локус). Гены расположенные в одном и том же локусе парных хромосом, называют аллелями. Каждый аллель представляет собой измененное состояние одного и того же гена.
Если оба аллельных гена одинаковы, то говорят, что кролик гомозиготен по данному признаку, если различны, то гетерозиготен. У гетерозиготных животных один ген может быть доминантным, т.е. подавлять действие другого (аллельного ему) гена. Доминантный ген обозначают большой буквой, а подавляемый ген (рецессивный) обозначают маленькой буквой.
Доминантный ген R и его рецессивный аллель r обусловливает длину и толщину остевых и переходных волос. При этом ген R ответствен за нормальную структуру волос, а r - за измененную характерную для кроликов типа рекс. Доминантный ген проявляет свое действие и когда находиться в одной из гомологичных хромосом (гетерозиготное состояние) и когда присутствует в обеих хромосомах (гомозиготное состояние).
Рецессивные гены обнаруживают свое присутствие только в гомозиготном состоянии, например rr - вызывает укорачивание ости и пуха, недоразвитие усов, характерные для коротковолосых кроликов типа рекс.
Понятие о гомо- и гетерозиготности, ключевое понятие в селекции, именно этим объясняется появлением у нормально волосных родителей типа Rr коротковолосых крольчат в помете. На основании законов Г.Мендаля можно рассчитать процентное соотношение в потомстве различных генотипов: Rr x Rr = RR(25%), rr(25%), Rr(50%). Внешне RR и Rr выглядет одинаково, а при скрещивании получим классическое соотношение по фенотипу 3:1.
Отличить RR и Rr можно только опытным путем. Такие проверочные скрещивания позволяют при необходимости избавиться от нежелательного рецессивного гена за 4-5 поколений с учетом качества потомства.
Понятие "доминантность" и "рецессивность" относиться не ко всему животному в целом, а только к тем его наследственным факторам, которые находятся в одной и той же паре хромосом и определяют, как правило, один признак. Например. фактор серой ("дикой") окраски обозначается символом G и являеться доминантным к g. Гетерозиготное животное Gg будет серым, так же как и Gg, а gg - черным.
Разведение между собой родственных животных увеличивает долю генов приведенных в гомозиготное состояние. И хотя получить гомозиготность по всем генам невозможно, длительно родственное разведение повышает однородность стада и снижает эффективность отбора. Полные братья и сестры(потомство одних родителей) имеют примерно 50% одинаковых генов. Степень генетического родства полубратьев и полусестер в 2 раза ниже, т.е. 25%. Такая же степень родства между дядей и тетей, племянниками, между дедушкой. бабушкой и внуками. Если родственные животные обладают ценными качествами, родственное разведение (инбридинг) закрепляет эти качества. Вместе с тем родственное разведение закрепляет и сходные недостатки. Нежелательные признаки чаще обусловлены рецессивными генами. Переводя нежелательные и вредные рецессивы в гомозигоное состояние, инбридинг делает скрытое явным. Коэффициент инбридинга (степень возрастания гомозиготности) - это вероятность, с какой ген, имеющийся у общего предка родителей, окажется в гомозиготном состоянии у потомства. При спаривании братьев и сестер в течение одного поколения коэффициент инбридинга равен 25%, двух поколений - 37,5%, трех поколений - 50%, десяти поколений - 88,6%. Т.о. даже после 10 поколений такого близкородственного скрещивания сохраняется значительная доля гетерозигоности. Только в сочетании со строгим отбором родственное разведение может обеспечить высокую однородность стада по желательным генам и признакам. Очень трудно избавиться от нежелательных рецессивных генов. Даже после 20 поколений отбора по фенотипу 3% особей будут носителями таких генов.
Совокупность всех наследственных задатков организма называют генотипом. Под влиянием внутренних причин и разнообразных внешних условий многие задатки реализуют себя не в полной мере. Наконец некоторые гены могут измениться в процессе развития - мутировать. Все это в той или иной мере отражается на фенотипе животного. Под фенотипом понимают совокупность признаков и свойств проявившихся в результате взаимодействия генотипа и среды, т.е. конкретного проявления признаков с которыми работает селекционер. (Ниже см. таблицу основных генов окраски)
Доминантный ген С определяет наличие пигмента, гены B, D, E в совокупности обуславливают развитие черного пигмента. ген А отвечает за его зональное распределение. Кролики гомозиготные по рецессивному гену с, - альбиносы, у них полностью отсутствует пигментация волосяного покрова.
Мутация гена B в ген b обуславливает коричневую окраску. Ген D - усилитель черного пигмента в корковом и сердцевинном слое волоса, а ген d - разбавитель черного пигмента, вызывающий голубую окраску. Ген Е переводит желтую окраску в черную, рецессивный ген е определяет желтую окраску. Ген А (ген зональности) образует различно окрашенные зоны по длине волоса и одновременно вызывает осветление брюха, внутренней стороны лап и хвоста. Когда ген А мутирует в ген а, то исчезает зональность волоса и кролики имеют равномерное распределение пигмента по всему волосу.
Другие гены в комбинации с пятью основными (CBDEA) дают новые цветовые вариации. Так ген Н определяет так называемую горностаевую окраску, ген v обусловливает белый волосяной покров и цвет глаз (венский белый). Ген W регулирует ширину светлого кольца у зонально окрашенного кролика.
В некоторых случаях один и тот же ген мутирует несколько раз, создавая серию множественных аллелей. Например, ген С по степени убывания доминантности образует следующую серию: C>cchi>cd>cm>c, где cchi - ген темной шиншилловой окраски; сd - ген светлой шиншилловой окраски; сm - ген окраски типа мардер. Серия множественных аллелей гена Е состоит из четырех пар генов: ED>E>ev>e; серия аллелей гена А - из трех пар: A>at>a. Мутация гена L в рецессивный l привела к появлению длинноволосых пуховых кроликов. Под действием полимерных генов P1p1, P2p2, P3p3 образуеться серебристая окраска; генов Y1y1, Y2y2 - оранжевая; ген En способствует развитию пятнистости, характерной для породы бабочка.
Селекционера всегда интересует генотип, но определить его по фенотипу с большой точностью, за исключением окраски, к сожалению, невозможно. И несмотря на это, вся селекция, проводимая человеком как раньше, так и теперь, основана на оценке животных по фенотипу. Было и остается незыблемым правило: "Лучшие генотипы нужно искать среди лучших фенотипов". Генотип Фенотип Порода, тип CCBBDDEEAA Серо-заячья (агути), зональная Дикий кролик, фландр, серый великан, бельгийский заяц CCBBDDEEaa Черная, голубая Аляска, виенчин, венский голубой, бевернский голубой CCBBddEEaa Коричневая Гавана CCbbDDEEaa Серо-голубая Белка магдебургская CCbbddEEAA Жемчужная Белка дюссельдорфская ccBBDDEEAA Белая (альбинос) Белый великан, новозеландская белая, белый бускат CCBBDDEEAAataty1y1y2y2 Черно-огненная Черно-огненный cHcHBBDDEEaa Белая с темными ушами, кончиками морды, лап и хвоста Русский горностаевый, калифорнийская CCBBDDeeaa Желтая неравномерная Тюрингский паломино cchicchiBBDDEEAA Серо-голубая, зональная Шиншилла cmcmBBDDEEaa Темно-коричневая Советский мардер CCBBDDEEaaEnEn Белая пятнистая Бабочка CCBBDDEDEDAA Черно-бурая, на боках зональная Черно-бурый CCBBDDevevAA Чередование черных, желтых и белых (на юрюхе) пятен, зональная Японский CCBBDDEEaaP1P1P2P2P3P3 Чередование пигментированных (черных) и непигментированных остевых волос, пух голубой Шампань, серебристый ccBBDDEEAAll Белая (альбинос), волос длинный Белая пуховая, ангорская CCBBDDEEaarr Черная, волос короткий Рекс черный CCBBddEEaarr Голубая, волос короткий Рекс голубой ccBBDDEEAArr Белая, волос короткий Рекс белый
Коэффициенты наследуемости. При отборе по фенотипу эффект достигается медленно, потому, что фенотипические различия не всегда соответствуют генотипическим. Такие хозяйственно-полезные признаки как скорость роста, молочность, многоплодие... в отличии от окраски в значительной степени зависят от внешних маскирующих факторов окружающей среды. Один и тот же признак может иметь разный коэффициент наследуемости (h2) в разных стадах. Вычисление коэффициента наследуемости в конкретном стаде позволяет предвидеть эффективность селекции по фенотипу. Коэффициент наследуемости показывает степень соответствия фенотипа генотипу.
Усредненные коэффициенты такие: 0,10-0,04 скорость роста от отъема (28дн.) до 70 дней 0,4-0,6 оплата корма за тот же период 0,3-0,4 оплата корма при постоянной скорости роста 0,3 масса тушки при постоянной живой массе 0,68 выход мышечной массы 0,39 выход жира 0,53 живая масса в 84 дня 0,39 живая масса в 42 дня 0,52 число крольчат при отъеме
Чем более "жестко запрограммирован" признак, тем выше его коэффициент наследуемости. Но это не означает, что признак с низким коэффициентом наследуемости не наследуются, для них не эффективен отбор по фенотипу.
Быстрота наследственного улучшения стада по конкретному признаку зависит от следующих основных факторов: интенсивности отбора (величины селекционного дифференциала); величины коэффициента наследуемости (h2); быстроты смены поколений (интервала между поколениями, выражаемого средним возрастом родителей в момент, когда рождено примерно 50% их потомства).
Нужно как можно точнее оценивать селекционные признаки - это увеличит наследуемость, не удлинит без особой необходимости интервал между поколениями, необходимо обеспечить оптимальную интенсивность отбора. Последнее возможно только в достаточно большом стаде, чтоб было из чего выбирать. Высокая плодовитость кроликов позволяет применять строгий отбор. Важно правильно определить признаки отбора - не слишком разбрасываться. Желая усилить один из признаков отбора мы неизбежно уменьшим интенсивность отбора по другим признакам. чем больше признаков отбора, тем менее строгий отбор по каждому отдельному признаку. Нередко между селекционными признаками существует отрицательная корреляция, т.е. по мере улудшения одного признака отрицательно с ним коррелирующий ухудшается. Все это требует глубокого изучения и выработки определенной тактики отбора.
Генетика окрасов кроликов
Окраска агути
Дикий кролик одет серым мехом, который сверху более тёмен. Нижняя часть туловища, около горла, живот и внутренняя сторона ног и низ хвоста окрашены в белый цвет. Характерная серая окраска шкурки дикого кролика зависит от зонарного распределения пигмента по длине волоса. Зонарное распределение пигмента состоит в том, что в волосе имеются 3 зоны: на конце волоса и у основания – чёрные, между ними – жёлтая.
Дикий окрас является приспособленческой окраской кролика. Появление в результате какой-либо мутации белой окраски повлекло бы очень быструю гибель такого животного. Другое дело в условиях одомашнивания, где любая новая окраска интересна для человека с точки зрения получения новой породы.
Два основных признака характеризуют окраску агути: то, что волос пигментирован и то, что этот пигмент распределяется по волосу тремя кольцами. Наличие пигментации в волосе определяется один геном, который получил обозначение А. Когда ген А в результате мутации превратился в свой рецессивный аллеломорф а, то обнаружилось, что кролики, гомозиготные по этому гену (аа), полностью белые и с непигментированными красными глазами.
Зонарное распределение пигмента по волосу так же определяется один геном, получившим обозначение G. Когда ген G в результате мутации превратился в свой рецессивный аллеломорф g, то обнаружилось, что кролики, гомозиготные по этому гену (gg), чисто-чёрные. Зонарность исчезла. Чёрный пигмент, бывший до сих пор в виде колец у основания и начала волоса, распространился теперь по всему волосу, и вся шкурка оказалось окрашенной в равномерно чёрный цвет.
Однако интенсивная чёрная окраска развивается только при действии особого гена, обозначаемого D. Когда была найдена мутация гена D в его рецессивный аллеломорф d, то обнаружилось, что кролики (dd) вместо интенсивного чёрного цвета окрашены в голубой цвет.
Окраска дикого кролика (агути) сложна по своей природе. Её определяет совокупность 5-ти генов. Ген А обуславливает наличие пигмента, гены B, C, D в совокупности обуславливают полное развитие чёрной окраски. Ген G даёт зонарную окраску волоса.
Окраска агути свойственна разводимым домашним породам – бельгийский великан (фландр), французский баран. Это наиболее старые по-своему происхождения породы. То мясное направление разведения кролика, что имело место в средние века, заставляло обращать главное внимание на величину кролика и его мясные качества, а не на его окраску.
Основные гены окраски кроликов A, B, C, D и G и их действие
Ген А – основной ген пигментации, способность кролика производить пигмент. Кролик, гомозиготный по рецессивному аллеломорфу гена А – аа – абсолютно не пигментирован, белый, причём глаза благодаря просвечиванию кровеносных сосудов становятся красными, т. е. животные становятся типичными альбиносами.
Ген В. Если мы получим животных, имеющих генотип AA bb CC DD gg, то увидим, что кролики оказываются окрашенными в жёлто-чёрный цвет (черепаховые кролики). Это показывает, что полное развитие чёрного пигмента происходит только у тех животных, у которых есть ген В (AA BB CC DD gg – кролик чёрного окраса, AA bb CC DD gg – кролик жёлтого-чёрного окраса). Ген В является усилителем пигментации и переводит жёлто-чёрную окраску в чисто чёрную. Кролик AA bb CC DD GG будет жёлтым агути с белым брюшком.
Ген С так же оказывается усилителем пигментации. Если мы получим животное гомозиготное по его рецессивному аллеломорфу, то есть животное с генотипом AA BB сс DD gg, то его окраску будет коричневой. Ген С усиливает пигментацию от коричневого до чёрного, в то время как ген В усиливает пигментацию от жёлто-чёрного до чёрного. Однако коричневым этот кролик будет лишь при отсутствии гена агути G (AA BB сс DD gg). Если кролик имеет генотип AA BB сс DD GG, то это уже будет коричневый агути с характерным зонообразным проявлением пигмента и белым брюшком.
Ген D. Этот ген является усилителем пигментации и обуславливает развитие чёрного пигмента в корковом и мякотном веществе волоса. Его рецессивный аллеломорф (d) обуславливает пигмент главным образом в мякотном веществе волоса. В корковом же веществе пигмент имеется в небольшом количестве. Чёрный пигмент, находясь в мякотном веществе, сквозь полупрозрачную среду коркового вещества придаёт волосу голубую окраску. Кролик, гомозиготный по рецессивному аллеломорфу (d), называется голубым (светлосерым) и имеет генотип AA BB CC dd gg. В комбинации с геном G мы получаем кролика окраса голубой агути.
Ген G. В случае гомозиготности по рецессивному аллеломорфу (g) гена G, животные с генотипом AA BB CC DD gg окрашены в чёрный цвет.
Первичные мутационные типы окраски
Благодаря мутационному изменению каждого из 5-ти генов окраски агути AA ВВ CC DD GG мы получаем 5 различных генотипов, которые называют первичными мутационными типами окраски:
aa BB CC DD GG - белый
AA bb CC DD GG – жёлтый агути
AA BB сс DD GG – коричневый агути
AA BB CC dd GG – голубой агути
AA BB CC DD gg – чёрный
Первоначально все породы разводимые человеком были окрашены в дикий цвет агути. Затем среди этих однородных животных стали появляться различные окраски как следствие мутации различных генов. Путём отбора эти признаки были закреплены и дали начало новым породам.
Первоначальные мутационные типы окраски присущи некоторым разводимым сейчас породам:
aa BB CC DD GG – Белый Великан, Польские кролики
AA BB CC DD gg – Аляска
Ген Х
Однако к 5-ти генам окраски надо добавить ещё один. Это ген Х, который так же должен быть назван основным геном пигментации. Только в его присутствии полностью развивается окраска в шкурке животного.
Действие гена Х сильно отличается от действия гена А. Животные, гомозиготные по гену аа, всегда полные альбиносы, то есть имеют белую шкурку и непигментированные красные глаза. Кролики, гомозиготные по гену хх, оказываются не полностью лишены способности вырабатывать пигмент. Они имеют белую шкурку, но пигмент остаётся в глазах – глаза окрашены в голубой цвет. Такие кролики получили название венских белых и их генотип AA xx BB CC DD GG.
Интересный результат получается при скрещивании двух белых кроликов: альбиноса aa XX BB CC DD GG и белого венского AA xx BB CC DD GG. Их скрещивание даёт окрашенных кроликов Aa Xx BB CC DD GG.
Комбинации генов окраски
При скрещивании друг с другом первичных мутационных генов окраски было получено значительное количество комбинаций, имеющих различные фенотипы. Получение различных комбинаций возможно только по генам B,C,D,G (они дают 16 различных комбинаций (животные не белые)). Животные же гомозиготные по рецессивному аллеломорфу гена А (аа) будут всегда белыми красноглазыми.
Первой комбинированной породой были, по-видимому, венские голубые, появившиеся в 1890-х годах. Исходными были 2 породы: бельгийский великан (агути - AA BB CC DD GG) и "Mahrishes Kaninchen” (голубой агути - AA BB CC dd GG). По видимому одно животное было гетерозиготным по гену агути.
Скрещивание этих пород имело вид:
Р (скрещивание) AA BB CC DD Gg х AA BB CC dd GG
F1 (первое поколение) AA BB CC Dd GG – агути гетерозиготный по одному гену
AA BB CC Dd Gg – агути гетерозиготный по двум генам
F2 (второе поколение от скрещивания AA BB CC Dd Gg х AA BB CC Dd Gg)
9 AA BB CC D G – серые кролики (агути)
3 AA BB CC D gg – чёрные кролики
3 AA BB CC dd G – кролики голубые агути
1 AA BB CC dd gg – голубые кролики
Таким образом, венские голубые были получены как новая комбинация, гомозиготная сразу по двум рецессивным генам - AA BB CC dd gg.
Имеются 2 породы венских голубых: одна – более тёмная (AA BB CC dd gg H), вторая – светло-голубая ("империал” - AA BB CC dd gg hh). Разница между ними определяется особым геном H, который является усилителем пигментации и его действие изучено на фоне гена d.
Подобным же образом была получена порода гавана, кролики которой окрашены в коричнево-шоколадный цвет. Её возникновение относится в 1890-ым годам, причём известно, что исходная самка была коричневым агути, в её помёте наряду с животными агути был обнаружен кролик полностью коричневый. Данные относительно отца отсутствуют, но очевидно, что он должен был быть или коричневым агути или чёрным, гетерозиготным по гену G. В результате скрещиваний были получены AA BB сс DD Gg, которые при скрещивании между собой дали:
AA BB сс DD GG + AA BB сс DD Gg + AA BB сс DD gg
1 2 1
AA BB сс DD gg – кролик окрашенный равномерно в коричневый цвет – гавана.
Таким же способом была получена комбинация гомозиготная по рецессивным генам – b и g (AA bb CC DD gg), окрашенная в чисто жёлтый цвет без агути, свойственная породе тюрингенский. Правда в противоположность "гаване” и голубому жёлтая окраска не получается однородной. На шкурке можно найти тёмные черноватые пятна, переходящие в голубовато-серые. Жёлтый цвет у тюрингенских кроликов сильно варьируется от ярко-жёлтого до более ослабленного, что по-видимому зависит от наличия ряда модификаторов этой окраски и кроме того от наличия генов D и C в гетерозиготном состоянии.
От скрещивания венского голубого кролика и кролика окраса "гавана” можно получить комбинацию, содержащую из 3 рецессивных гена (cdg). Такая порода была получена в Марбурге в 1916 году. Окраска таких кроликов оказывается светло-голубоватой.
AA BB сс dd gg – марбургская белка.
Но существует 2 породы "белка”, различные по своему генотипу. У дюссельдорфской белки есть зонарное распределение пигмента по волосу отчего светло-голубая окраска приобретает несколько грязноватый оттенок. Животное как и все агути имеет белое брюшко.
AA BB сс dd GG – дюссельдорфская белка.
От скрещивания тюрингена с "гаваной” мы получаем комбинацию трёх рецессивных генов (AA bb cc DD gg) .
Р (скрещивание) AA bb CC DD gg x AA BB сс DD gg
F1 (первое поколение) AA Bb Cc DD gg – чёрный гетерозиготный по 2-ум генам
F2 (второе поколение от скрещивания AA Bb Cc DD gg х AA Bb Cc DD gg)
9AA B C DD gg – чёрные кролики
3AA B сс DD gg – кролики окраса "гавана”
3AA bb C DD gg – тюрингенские кролики
1AA bb cc DD gg – оранжевый кролик
При скрещивании оранжевого кролика (AA bb cc DD gg) с голубым (AA BB CC dd gg) можно получить комбинацию 4-х рецессивных генов (AA bb cc dd gg):
Р (скрещивание) AA bb cc DD gg x AA BB CC dd gg
F1 (первое поколение) AA Bb Cc Dd gg – чёрный гетерозиготный по 3-ём генам
F2 (второе поколение от скрещивания AA Bb Cc Dd gg х AA Bb Cc Dd gg)
27AA B C D gg – чёрные кролики
9AA bb C D gg – тюрингенские кролики
9AA B сс D gg – кролики окраса "гавана”
9AA B C dd gg – голубые кролики
3AA bb сс D gg – оранжевые кролики
3AA bb C dd gg – кролики светлые тюрингены
3AAB cc dd gg – кролики окраса белка марбургская
1AA bb cc dd gg – кролик песочного окраса
Кролики песочного окраса являются прекрасными анализаторами при проверке производителей вследствие своей рецессивности по 4-ём генам.
Так же в результате тригибридного расщепления был получен ещё один новый генотип - AA bb CC dd gg. Эти кролики окрашены в светло-черепаховый цвет.
Итак, мы получили 16 комбинаций изученных 4-х пар генов (B-b, C-c, D-d, G-g):
1) AA BB CC DD GG – агути ("Бельгийский великан”, дикий кролик)
2) AA BB CC DD gg – чёрный ("Аляска”)
3) AA BB CC dd GG – голубой агути
4) AA BB cc DD GG – коричневый агути
5) AA bb CC DD GG – жёлтый агути
6) AA BB CC dd gg – голубой ("Венский голубой”)
7) AA BB cc DD gg – коричневый ("Гавана”)
8) AA bb CC DD gg – жёлтый ("Тюрингенский”)
9) AA BB cc dd GG – светло-голубой агути ("Белка дюссельдорфская”)
10) AA BB cc dd gg – светло-голубой ("Белка марбургская”)
11) AA bb CC dd GG – светло-черепаховый агути
12) AA bb CC dd gg – светло-черепаховый
13) AA bb cc DD GG – оранжевый агути
14) AA bb cc DD gg - оранжевый
15) AA bb cc dd GG – песочный агути
16) AA bb cc dd gg – песочный
Серия множественных аллеломорфов некоторых генов окраски (A, B и G)
Каждый из 5-ти генов окраски способен мутационно изменяться на новый ген: ген В имеет мутацию виде рецессивного аллеломорфа b, ген А мутировал в а, ген С - в с, ген D – в d, ген G – в g. Что касается генов C и D, то известно, что у них нет других аллеломорф кроме вышеуказанных.
Для генов A, B, G известны ряды аллеломорфов, которые произошли последовательно друг от друга. Каждый аллеломорф обуславливает свою особую, характерную для него окраску.
Серия аллеломорфов гена А слагается из шести генов, гена В – из пяти, гена G – их трёх. Почти все новые аллеломорфы являются рецессивными по отношению к исходным генам, за исключением трёх доминантным над геном B.
Серия множественных аллеломорфов гена А
Серия множественных аллеломорфов гена А состоит из 6-ти генов. Ген А является основным геном пигментации. Ген achi определяет красивую светло-серую окраску так называемого кролика шиншилла. Ген ad определяет также окраску шиншиллы, но более светлую. Ген am обуславливает развитие шоколадно-коричневой окраски с некоторыми частями тела окрашенными в более тёмный цвет. Такая окраска называется "мардер”. Ген аn определяет развитие характерной расцветки называемой горностаевой, русской или гималайской. Ген а – ген альбинизма, при котором пигмент не развивается.
Ген achi появился впервые в 1910 году во Франции. В 1921 году было доказано, что ген шиншилла является аллеломорфом основного гена цветности А. Ген аchi обуславливает некоторое (очень небольшое) осветление интенсивности чёрного пигмента и кроме того ликвидирует жёлтый пигмент шкурки. Окраска шиншилла возникает благодаря взаимодействию гена achi и гена зонарности G. Волос у шиншиллы обладает зонами – чёрной, белой и чёрной. Генная формула – achi achi BB CC DD GG.
Совсем по-иному выглядят кролики гомозиготные по гену achi, у которых ген G имеет вид g - achi achi BB CC DD gg – эти кролики очень сходны с обыкновенными чёрными кроликами, но отличаются от них отсутствием блеска, свойственно чисто-чёрным кроликам. Это окрас "чёрная сепия”.
Ген ad – ген светлой шиншиллы – впервые появился в Америке в 1925 году. В 1926 году было доказано, что данный ген является членом серии аллеломорфов гена А. Ген ad ликвидирует жёлтый пигмент, но настолько сильно что уже затрагивает и чёрный. Кролик ad ad BB CC DD gg окрашен в чёрный цвет меньшей интенсивности. Таких кроликов называют "тёмная сепия”. Если на фоне тёмной сепии начинает работать ген G, то кролик имеет окраску сходную с шиншиллой. Волос так же имеет 3 зоны: чёрную, белую и чёрную. Однако так как чёрный пигмент у этих кроликов так ослаблен, то окраска этого кролика будет более светлая, чем у обыкновенной шиншиллы. Окрас ad ad BB CC DD GG называется светлой шиншиллой.
Просмотров:
347
|
Добавил:
k-9
|
Дата:
05.04.2018
|
Ген тёмной шиншиллы не полностью доминантен по отношению к гену светлой шиншиллы, поэтому гетерозиготные животные achi ad BB CC DD GG имеют более светлую шкурку, чем achi achi BB CC DD GG.
Ген am ликвидирует жёлтый пигмент и в её большей степени влияет на чёрный. Ген am обуславливает характерную кунью окраску, благодаря которой эти кролики названы "мардер” (куница). Впервые мардеры были обнаружены в 1925 году в Германии, где возникли как мутация гена achi. Выставочный "мардер”, представляющие собой желаемый образец породы, имеют по всей поверхности туловища коричневый пигмент, особенно интенсивный на спине, где он образует широкую каштанового цвета полосу, хорошо отличающуюся от более светлых боков. Конечности, мордочка и уши более сильно пигментированы, чем всё остальное туловище и имеют коричнево-чёрный цвет. В ирисе глаза имеется коричневый пигмент, но здесь образование пигмента в противоположность полностью окрашенному кролику сильно редуцировано, и можно уже заметить лёгкое просвечивание кровеносных сосудов, что даёт в темноте красноватый отблеск в глазах.
Эта ценная кунья окраска, свойственная породе "мардер”, определяется гетерозиготностью гена am по одному из более рецессивных генов той же серии. Гомозиготный мардер (am am) оказался почти чёрным животным и был назван "светлой сепией”. Глаза гомозиготного мардера отличаются от глаз гетерозиготного мардера так как не дают того ясного красноватого отблеска, который имеется у последнего.
Типичный мардер является геторозиготным и имеет генотип am an BB CC DD gg. В случае гетерозиготности его по гену а (ген альбинизма) наблюдается его очень сильное осветление причём отметины головы и конечностей очень сильно выделяются. Вся окраска почти светло-коричневая. Всё это показывает, что разводить ценную породу мардер в чистом виде невозможно.
Скрестив двух типичных мардеров, мы всегда обнаружим расщепление в отношении 1 : 2 : 1 :
P am an BB CC DD gg x am an BB CC DD gg
F1 am am BB CC DD gg + 2 am an BB CC DD gg + an an BB CC DD gg
тёмные мардеры мардеры горностаевые
Для того чтобы получить мардеров желаемой окраски надо скрещивать между собой тёмного мардера и горностаевого кролика:
P am am BB CC DD gg x an an BB CC DD gg
F1 am an BB CC DD gg
мардеры
Окраска мардер имеется только у кроликов, у которых нет зонарности (G). В том случае, если мы имеем кроликов с генотипом am am BB CC DD GG, то их фенотип очень сходен с окраской шиншиллы. Ген am уничтожает жёлтый пигмент и сильно осветляет чёрный цвет. При этом зонарность имеет вид: чёрное кольцо, белое кольцо, чёрное кольцо. Поэтому кролик генотипа am am BB CC DD GG (зонарный мардер) очень сильно похож на кролика шиншиллу, но имеет буро-коричневые пятна на нижней части морды, в области нижней челюсти, по бокам туловища и иногда на ушах.
Ген an определяет расцветку горностаевого кролика. Имеет несколько названий – русский, горностаевый, гималайский. Возник он как мутация гена А среди кроликов дикой окраски. Окраска кролика чисто белая, нос, уши, лапки, хвостик пигментированы. Глаза у горностаевого кролика лишены пигмента, в силу этого просвечивание кровеносных сосудов делает их красными. Таким образом, мы видим переход от полностью окрашенных глаз у кроликов агути и шиншилла к глазам мардера, у которого пигментация глаз ослаблена и они имеют красноватый отблеск, и к глазам горностаевого кролика, полностью лишенных пигмента.
Горностаевые кролик рождаются полностью белыми, и лишь к 2-3 неделям у них появляется пигмент на соответствующих местах. К 4-6-ти месячному возрасту их пигментация достигает полной интенсивности. Генотип гималайского кролика an an BB CC DD gg.
При наличии зонарности an an BB CC DD GG ген an уничтожает жёлтый пигмент и сильно осветляет чёрный цвет. При этом зонарность имеет вид: чёрное кольцо, белое кольцо, чёрное кольцо. Шкурка кролика имеет серебристый вид. Наличие гена G вместе с тем обуславливает наличие белого живота и белой нижней части хвоста.
Шестой член серии гена А – ген альбинизма (а). Это чисто белые животные с красными глазами (разводимые породы – "польские кролики” и "белые великаны” (1932 год)). В генотипе альбиноса может находиться любой набор генов окраски.
1)AA BB CC DD GG – агути ("Бельгийский великан”, дикий кролик)
2)achi achi BB CC DD GG – шиншилла тёмная (Тёмная шиншилла)
3)ad ad BB CC DD GG – шиншилла светлая (Светлая шиншилла)
4)am am BB CC DD GG – шиншилла светлая с коричневым оттенком
5)an an BB CC DD GG – горностаевый агути
6)aa BB CC DD GG – альбинос ("польские кролики”, "белый великан”)
7)AA BB CC DD gg – чёрный ("аляска”)
8)achi achi BB CC DD gg – сепия чёрная
9)ad ad BB CC DD gg – сепия тёмная
10)am am BB CC DD gg – сепия светлая
11)an an BB CC DD gg – горностаевый ("русский” или "горностаевый”)
12)aa BB CC DD gg – альбинос ("польские кролики”, "белый великан”)
Взаимодействие генов серии А с другими генами
Каждый из генов серии А может быть комбинирован с другими генами окраски. Так как гены серии А редуцируют жёлтый пигмент, то интересны их комбинации с генами, определяющими этот жёлтый пигмент. Разберём комбинации членов серии гена А с геном b.
AA bb CC DD gg – жёлтый кролик (жёлто-чёрный). Можно заранее предсказать как будет окрашено животное гомозиготное по гену achi и по гену b – achi achi bb CC DD gg. Так как благодаря гену b может проявиться желтый пигмент, то животное, казалось бы, должно быть окрашено в жёлтый цвет. Но имеющийся ген achi полностью ликвидирует жёлтый пигмент и, стало быть, животное должно быть лишено пигмента и оно должно белое.
Справедливость этого утверждения была доказана Кастлем и была получена данная комбинация. Это были чисто белые животные с голубыми глазами. Фенотип этих животных был схож с фенотипом белых венских AA xx BB CC DD GG. Полное различие их генотипов доказывается при их скрещивании. В результате получились животные, окрашенные в цвет агути.
P achi achi bb CC DD GG x AA xx BB CC DD GG
F1 A achi Хх Bb CC DD GG
агути
Получение белых животных генотипа achi achi bb CC DD GG осуществляется с помощью простого дигибридного расщепления (шиншилла х жёлтый агути):
P achi achi BB CC DD GG х AA bb CC DD GG
F1 A a chi Bb CC DD GG
агути
F2 9 A B CC DD GG – агути
3 A bb CC DD GG – жёлтый агути
3 achi achi B CC DD GG – шиншилла
1 achi achi bb CC DD GG – белые с голубыми глазами
Вслед за Кастлем Коссвинг также получил жёлтую шиншиллу, которая отличалась от кроликов Кастля. Во-первых, на кончиках волос они несли слабые следу пигментации, что придавало животным слегка грязноватый вид. Во-вторых, глаза животных были не голубые, а коричневые. Первое различие можно отнести за счёт того, что Кастль использовал в своих скрещиваниях не кроликов achi, а кроликов ad. Что касается окраски глаз, то кролики окраса шиншилла встречаются как с коричневыми, так и с голубыми глазами. Возможно, что окраска глаз зависит от самостоятельного гена. Есть указания, что коричневая расцветка глаз является доминантной по отношению к голубой.
У кролика с генотипом achi achi bb CC DD gg имеется пигментация на некоторых участках шкурки. Фенотип этого кролика похож на плохо окрашенного горностаевого кролика. Сходный фенотип имеет и комбинация гена am (мардер) c b и g - am am bb CC DD gg. Комбинация an an bb CC DD gg имеет очень ослабленную пигментацию.
Были так же получены комбинация серии генов А с генами с и d:
achi achi BB cc DD GG – коричневый агути.
achi achi BB cc DD gg – похож на обычную "гавану”.
achi achi BB СС dd GG – очень редкая комбинация "голубая белка” (blue sguirrel).
Получены так же комбинации an с d, c и g:
an an BB CC dd GG – голубой агути горностаевый
an an BB CC dd gg – голубой горностаевый
an an BB cc DD GG – коричневый агути горностаевый
an an BB cc DD gg – коричневый горностаевый
1)AA bb CC DD GG – жёлтый агути
2) achi achi bb CC DD GG – грязно-белый с голубыми или коричневыми глазами
3) ad ad bb CC DD GG – белый с голубыми или коричневыми глазами
4) am am bb CC DD GG - не разводилась
5) an an bb CC DD GG - не разводилась
6) aa bb CC DD GG – альбинос ("польские кролики”, "белый великан”)
7)AA bb CC DD gg – жёлтый (тюрингенский)
8) achi achi bb CC DD gg – грязно-белый с отметками русского кролика
9) ad ad bb CC DD gg – то же
10) am am bb CC DD gg – то же
11) an an bb CC DD gg – жёлтый русский
12) aa bb CC DD gg – альбинос ("польские кролики”, "белый великан”)
Просмотров:
324
|
Добавил:
k-9
|
Дата:
05.04.2018
|
ОКРАСЫ В ПОРОДЕ КИТАЙСКИЙ ШАРПЕЙ
ГЕНЕТИКА ОКРАСОВ У ШАРПЕЕВ
ЭТА СТАТЬЯ ПОСВЯЩЕНА МНОГООБРАЗИЮ ОКРАСОВ В ПОРОДЕ ШАРПЕЙ. КАКИЕ ОКРАСЫ ПРИНЯТЫ ПО СТАНДАРТУ. КАКИЕ СЧИТАЮТСЯ РЕДКИМИ. КАК МОЖНО ПОЛУЧИТЬ ЩЕНКОВ ОПРЕДЕЛЕННОГО ОКРАСА. И КАК ОКРАСЫ ЩЕНКОВ ЗАВИСЯТ ОТ ОКРАСОВ ИХ РОДИТЕЛЕЙ. ГЕНЕТИКА ОКРАСОВ ПОРОДЫ ШАРПЕЙ - ЭТО ИНТЕРЕСНО. АВТОР ЭТОЙ УНИКАЛЬНОЙ СТАТЬИ - ЕЛЕНА ИВАЩЕНКО.
ПОДРОБНОЕ ОПИСАНИЕ ОКРАСОВ ПОРОДЫ ШАРПЕЙ
Стандартом FCI (FCI-Standart #309 14.04.1999) породы Китайский шар-пей допускается любой сплошной окрас, исключая чисто белый (альбинос). Буквально это означает, что любой шар-пей, не имеющий пятен, разводов, подпалов и чепрака, вписывается в стандарт по окрасу. Однако в реальности до недавнего времени далеко не все сплошные окрасы можно было регистрировать официально. В реестре окрасов не было таких редких окрасов как изабелла, лиловый, изабелла-дильют. Поэтому заводчики были вынуждены регистрировать их, "подгоняя" под наиболее близкие по внешнему виду окрасы. Так, изабеллу регистрировали как олений, кремовый, а лиловый как шоколад и голубой. Изабелла-дильют регистрировался как абрикосовый дильют или крем-дильют. Надо ли говорить, что подобные регистрации только добавляли путаницы в родословных, становились причиной появления "артефактов" в разведении и давали повод для неверных выводов начинающим разведенцам породы.
В конце 2005 года на официальном уровне наконец-то была предпринята попытка систематизации окрасов в нашей породе. Результатом этой работы стал новый реестр окрасов, официально одобренный племенной комиссией РКФ. Регистрация окрасов собак породы Китайский шар-пей отныне должна производиться в соответствии с этим новым реестром.
Ниже приводится перечень принятых названий окрасов, описание фенотипических признаков. Так же в помощь заводчикам дана базовая генетическая расшифровка окрасов шар-пеев и основные принципы их получения.
I. РЕЕСТР РЕГИСТРИРУЕМЫХ РКФ ОКРАСОВ (ДЛЯ ПОРОДЫ КИТАЙСКИЙ ШАР-ПЕЙ)
С 2005 года регистрации подлежат следующие окрасы (даны точные названия):
Черный
Олений
Красный
Красный олень
Кремовый
Соболиный
Голубой
Изабелловый
Шоколадный дильют
Абрикосовый дильют
Красный дильют
Кремовый дильют
Лиловый
Изабелловый дильют
За рамками этого официального списка пока остается окрас "соболиный дильют", несмотря на то, что фактически он существует и является дильютной производной от соболиного окраса. Трудности с легализацией этого окраса в том, что даже соболиный окрас не всегда точно идентифицируется заводчиками и экспертами. Поэтому идентификация соболиного дильюта на данном этапе была признана еще более затруднительной. Заводчикам, у кого рождаются щенки, которых можно идентифицировать как соболиный дильют, пока по-прежнему будут вынуждены регистрировать их как абрикосовый дильют (данный окрас наиболее близок фенотипически к соболиному дильюту).
II. ФЕНОТИП. ОПИСАНИЕ ВНЕШНЕГО ВИДА СТАНДАРТНЫХ ОКРАСОВ В ПОРОДЕ КИТАЙСКИЙ ШАР-ПЕЙ.
Фенотип - это любой поддающийся наблюдению признак организма. Он формируется под влиянием генотипа (индивидуального набора генов), но не является внешним повторением генотипа, т.к. один и тот же фенотип может быть результатом разного набора доминантных и рецессивных генов.
Окрас, каким мы его видим невооруженным глазом – это фенотип. К примеру, перед нами стоят две черных собаки. Черный окрас – это то, что мы видим. При этом мы знаем, что первая собака получена от красного и шоколадного родителей, а вторая собака – от голубого и кремового. Опираясь на эти знания, мы говорим, что первая черная собака является носителем красного и шоколадного генов, а вторая – носителем голубого и кремового. Утверждая так, мы говорим о генотипах этих собак, и понимаем, что они разные. Но внешне перед нами две совершенно одинаковые по окрасу собаки.
Другой пример: перед нами две собаки, окрас которых мы идентифицируем как абрикосовый дильют. При этом у одной собаки имеется шоколадная мочка носа, а у другой нос практически розовый. В вязках с красными масковыми собаками первая будет давать красных и оленьих щенков (и, возможно, дильютных), а вторая собака в такой же комбинации может дать часть красных (оленьих) и часть черных щенков (или даже полностью черный помет). В данном случае мы так же имеем двух абрикосовых дильютных собак с совершенно разными генотипами, выявить которые можно только опытным путем (в определенных вязках). О генотипах речь пойдет позже. А пока опишем фенотипы, т.е. внешний вид собак каждого стандартного окраса.
При этом для удобства описания сразу разделим все окрасы на "основные", т.е. имеющие черный/шиферный пигмент, и "дильютные", т.е. имеющие коричневый пигмент, или вообще лишенные темного пигмента.
ОСНОВНЫЕ ОКРАСЫ:
Черный. Доминирующий окрас у шар-пеев. Может иметь оттенки иссиня-черного (очень редко встречается у взрослых шар-пеев, даже если они таковыми рождаются), очень темного коричневого, оливкового, а так же бурый отлив. Многие черные шар-пеи имеют "грязный" черный окрас. Как правило, такие собаки рождены от красных (оленьих) и шоколадных родителей, либо хотя бы один их родителей был не черный. Некоторые оттенки черного окраса заводчики между собой называют "серебристым соболем". Идеально черный окрас у шар-пеев – большая редкость. Порой из-за присутствия коричневого или синего оттенка у черных щенков заводчики называют их "коричневыми" или "голубыми". Рекомендуем не вводить в заблуждение ни себя, ни будущих владельцев этих щенков, и регистрировать их как черных (каковыми они и являются фенотипически и генотипически, несмотря на наличие отлива шерсти).
Олений. Вероятно, это один из самых древних окрасов шар-пея, т.к. название "собака с песочной шерстью" (на китайском примерно так: "Ша-пи до") могло относиться не только к фактуре шерсти, но и к ее цвету. Вариации этого окраса - оттенки золотистого, песочного. От светлого до довольно насыщенного, но без красноты. Особым отличительным признаком оленьего окраса является переход от более темного к более светлому оттенку (иногда значительно более светлому) в таких местах как внутренние стороны бедер, "зеркало" вокруг ануса, внутренние стороны передних лап, низ живота и груди. В то же время, большинство оленьих шар-пеев с типом шерсти "хорс" могут иметь достаточно равномерный светлый олений окрас. Зачастую в быту олений окрас называют "палевым", "песочным" или "рыжим". Подобные названия оленьего окраса до сих пор еще можно встретить и в официальных родословных некоторых шар-пеев. Однако регистрации подлежит название "олений", т.к. под "палевым" и "песочным" так же можно понять и кремовый, а под "рыжим" вообще все вариации окрасов – красный, олений, изабелла, и все их дильютные производные.
Красный. Один из самых распространенных окрасов среди современных шар-пеев. Вариации от насыщенного красного (как у ирландского сеттера) до светло-рыжего. Главный элемент окраса - рыжина, которая отсутствует (или слабо выражена) у собак оленьего окраса. Наиболее модным считается т.н. махагоновый окрас - очень яркий, сочный красный, как у ирландского сеттера. Несмотря на то, что этот окрас заводчики уже давно выделяют как вполне самостоятельную единицу, в официальных документах он регистрируется как "красный". Другое название красного окраса, которое можно услышать в быту и увидеть в некоторых родословных - "рыжий". Это не совсем корректно, т.к. "рыжий" - очень объемное обозначение, которое может включать в себя и олений, и красный окрасы, и их дильютные производные. Красный окрас – один из наиболее однородных светлых окрасов у шар-пеев. В отличие от оленьего окраса (и от красно-оленьего) красный окрас практически лишен переходов от темного к светлому, не имеет осветленных участков на бедрах, животе и т.д..
Красный олень. Данный окрас выделен в самостоятельный исключительно для удобства заводчиков, которые могут испытывать трудности с идентификацией окрасов у щенков в момент их актировки. Фактически, красный олень – это светло-красный не насыщенный окрас, может быть довольно "пестрым", т.е. иметь основные признаки оленьего окраса (белесые места под хвостом, на животе и т.д.), но в то же время быть более рыжим, чем олений окрас. Поскольку интенсивность окраса и оттенок (рыжий или песочный) у некоторых щенков становится очевидным только после первой линьки, не исключены ошибки в идентификации окраса при регистрации помета (в 45 дней). Если у заводчика есть сомнения, как правильно записать окрас – красным или оленьим, имеет смысл записывать его как "красный олень". У владельца щенка при желании всегда будет возможность сделать корректировку окраса, получив описание от эксперта-породника на одной из монопородных выставок.
Кремовый. Вариации от почти белого до цвета вареной сгущенки. Темные оттенки кремового иногда называют "абрикосовым" окрасом, что не совсем верно. Абрикосовый окрас - это дильютная производная от оленьего, о чем будет сказано ниже. Иногда неопытные заводчики ошибочно именуют кремовый окрас белым. Белый окрас в шар-пеях не допустим. Кроме того, у кремового шар-пея имеется более темный (песочный, коричневатый) шерстяной покров на спине, хвосте и, в особенности, - на ушах. Наличие маски или зачернение вокруг носа обязательно. В силу особенностей генотипа, большинство кремовых собак имеют ослабленную пигментацию мочки носа и других традиционно "черных" мест на теле собаки основного окраса. Слабость черного пигмента мочки носа носит название "слизанный" или "снежный" нос – это розовая или сильно осветленная мочка носа на фоне темной маски (зачернения). Далеко не все эксперты любят подобные носы, однако кремовых шар-пеев с хорошо пигментированным черным носом крайне мало. Поскольку, как правило, кремовый окрас неизбежно, благодаря особенностям генотипа, влечет за собой "слизанный нос", опытные эксперты-породники должны знать о такой специфике собак кремового окраса и не снижать за это оценку в ринге. Также у кремовых шар-пеев нередко недостаток черного пигмента выражается в не полностью прокрашенном языке (который должен быть темно-фиолетовым). Данный признак уже является нежелательным даже для кремовых собак.
Соболиный. Самый спорный окрас в нашей породе. Зачастую сами эксперты не уверены, какая собака стоит перед ними – соболиная или чепрачная. Но в генетике этот окрас имеет совершенно четкое обозначение, о чем мы поговорим позже. В научной литературе этот окрас имеет название "зонарный серый". У шар-пеев оно трансформируется в "соболиный". Внешне этот окрас выглядит как серое напыление по корпусу шар-пея, иногда с заходом на морду и частично на лапы. Так как зонарными являются только остевые волосы, густота серого напыления интенсивнее там, где скопления остевых волос больше – спина, хвост. На лапах ниже локтевых и скакательных суставов серый налет отсутствует. Отличие от чепрачного окраса принципиально – остевой волос у соболиного окраса – не сплошной черный, а зонарный, очень светлый у корня, рыжий посредине и черный на конце. Длина зон варьирует, часто рыжая зона (средняя) бывает короче двух других, и это дает более насыщенный соболиный окрас. Если рыжая зона длинее, то и соболиный окрас менее интенсивный.
Голубой. От нежного дымчато-серого до темно-серого, шиферного окраса шерсти. Мочка носа у светлых голубых собак обычно темнее. У темно-голубых – сливается с общим фоном. Не существует отдельных разновидностей голубого окраса, таких как "голубой" и "голубой дильют". Для всех голубых шар-пеев следует применять одно название – голубой, поскольку это собаки, имеющие один генотип, а слово "дильют" предполагает наличие качественных различий в генотипе, о которых мы поговорим ниже. Разведенцам нужно помнить, что голубой окрас - это ослабленный черный, и так же как и черный, относится к группе пигментированных. Производство черного пигмента по-прежнему происходит, но в меньшем объеме и гранулы пигмента в волосе расположены прерывисто, островками, поэтому визуально этот окрас смотрится как голубой - от темного до очень светлого. Вот простая аналогия для понимания того, что представляет собой голубой окрас: если вы возьмете черную краску, и добавите к ней воды, то получите определенный оттенок серого, интенсивность которого будет зависеть от количества воды в краске. Но суть остается одна - это по-прежнему черная краска. А вот у дильютных собак черная "краска" отсутствует в принципе. Пасть и язык голубых шар-пеев - темно-лавандовые, либо темно-фиолетовые, в зависимости от "разбавленности" черного пигмента. Светлый лавандовый язык у голубого шар-пея следует рассматривать как недостаток пигментации, т.к. фактически ген, разбавляющий черный окрас до голубого, почти не влияет на пигментацию языка, мочки носа, подушек лап и т.п. "деталей". Кожа тоже синеватая. Голубой - это один из самых модных окрасов и сравнительно молодой, т.к. первый голубой шар-пей родился в конце 1985 года. Погоня за закреплением окраса в породе сыграла плохую шутку с экстерьером голубых собак. Шоу-собаки среди шар-пеев этого окраса встречаются очень редко, и на выставках голубые собаки, как правило, сильно проигрывают шар-пеям других, в особенности, классических окрасов. Наиболее распространенные проблемы экстерьера голубых шар-пеев: большие уши, легкий костяк, тонкие ноги, маленькая голова, часто зауженная морда, узкая грудная клетка, пятнистый язык.
Изабелловый. Розовато-кремовый с голубым или серым отливом окрас. Производная голубых и "рыжих" (красных и оленьих) окрасов. При поверхностном взгляде может быть классифицирована как олений, светлый красный или даже темный кремовый, однако у собак этого окраса присутствуют атрибуты голубых собак: серый или шиферный нос, такая же явно серая, а не черная, маска на морде. Наличие этой темно-синей маски или серо-голубого напыления на морде обязательно. Кроме того в отличие от кремовых, красных и оленьих окрасов, шерсть изабеллы имеет синеватый, серебристый или холодный грязно-розовый отлив, без рыжего. Возможен более темный, сероватый (синеватый) или розовато-серый ремень на спине и затемнения на ушах. Опытные заводчики легко выделяют изабеллу в новорожденных пометах, особенно, если в них есть еще и оленьи или красные щенки. Новорожденная изабелла может быть серебристо-абрикосовой, серебристо-розовой, и даже серебристо-белой. Неопытный заводчик может принять за изабеллу новорожденного оленьего щенка, т.к. часто они рождаются почти серыми. О том, в каких случаях рождение изабеллы возможно, а в каких – нет, будет рассказано ниже.
Дильютные окрасы:
Шоколадный дильют. Название окраса говорит само за себя. Это шоколадный окрас, варьирующий в пределах от очень светлого "молочного" шоколада до довольно темного, насыщенного "горького" шоколада. Язык – лавандовый, подушки лап, мочка носа, анус – шоколадные. Если вам кажется, что перед вами – шоколадный шар-пей, но вы не уверены в этом, взгляните на цвет глаз и языка. Радужка глаз у шоколадного шар-пея – от темно-желтой до светло-коричневой, а язык – светло-сиреневый. Если ваш "шоколадный" шар-пей имеет синий язык – это черный окрас, имеющий коричневый оттенок. Фактически, название окраса "шоколадный дильют" – это "масло масляное", т.к. недильютным шоколадный окрас не бывает. В других породах собак для обозначения этого же окраса, используется термин "коричневый", но генетически это – тот же самый окрас.
Красный дильют. Красным дильютным окрасом принято называть любой красный окрас, не имеющий черного пигмента. При этом на генетическом уровне красный дильютный окрас может быть получен двумя совершенно разными способами, о которых речь пойдет в следующей главе. Следует знать, что красная дильютная собака может иметь маску на морде, но эта маска будет шоколадного окраса, а не черного или серого. Разница будет очевидна при сравнении со стандартной масковой собакой красного окраса. Точно так же красная дильютная собака может иметь шоколадный нос, шоколадные подушечки лап и анус. При этом язык у такой собаки будет светлым лавандовым, как у шоколадной дильютной собаки. При определенной комбинации генов, красная дильютная собака выглядит совершенно однотонной, без малейшего намека на шоколадный налет на морде, подушках, анусе. При этом у нее довольно темный лавандовый язык, что часто принимается за очень хорошую пигментацию. О генетических нюансах такого окраса пойдет речь далее.
Абрикосовый дильют. Данный окрас так же можно было бы назвать "оленьим дильютом", т.к. является его производной. Внешне – это светлый теплый золотисто-рыжий, именно, что "абрикосовый" окрас. Как и олений окрас, может иметь переходы от более темного на спине и голове, к более светлому на боках, животе и внутренних сторонах лап. По аналогии с красным дильютным окрасом, может иметь шоколадную маску, шоколадный нос или быть совершенно однотонным, без намека на шоколадный пигмент ("розовоносый" абрикосовый дильют). Трудности с этим окрасом в идентификации и в разведении точно такие же, как и с красным дильютным окрасом. Точно так же существует два варианта возникновения этого окраса, которые будут описаны позже.
Кремовый дильют. То же, что и кремовый окрас, вариации от почти белого до цвета вареной сгущенки, но без черного пигмента. И, так же как абрикосовый дильют и красный дильют, может быть двух генетических типов. Фенотипически первый проявляется в наличии сильно размытого светлого шоколадного пигмента, а второй выглядит почти белым, с фактически розовым носом, но хорошо пигментированным темно-лавандовым языком.
Просмотров:
350
|
Добавил:
k-9
|
Дата:
05.04.2018
|
Лиловый. Окрас, хорошо известный в породе Веймаранер. У шар-пеев до сих пор считается редким, хотя сейчас лиловых шар-пеев становится все больше. Вариации от розово-шоколадного до "теплого" розово-голубого или дымчатого серебристо-розового. При этом есть случаи рождения и очень темных лиловых шар-пеев, которых неопытные заводчики могут классифицировать как шоколадный дильют, однако все они имеют "сизый" налет на морде, невозможный у шоколадных собак. При сравнении со стандартным шоколадным окрасом разница становится очевидной. Маска не визуализируется (в тон окрасу), мочка носа - в тон окрасу.
Изабелловый дильют. Самый рецессивный окрас у шар-пеев. Серовато-розовый, в разных вариациях, от светлого до насыщенного, но без темного пигмента. При этом, так же как у абрикосового и красного дильюта бывает шоколадная маска, у изабеллового дильюта может присутствовать ЛИЛОВАЯ маска и лиловый ("сизый") нос.
III. Генотип. Описание генетических кодов стандартных окрасов в породе Китайский шар-пей.
Генотип – это наследственная конституция организма, индивидуальная совокупность всех его генов, которая контролирует развитие внешних признаков организма, т.е. его фенотип. Окрас, как мы его видим, может рассказать нам только о том, какие доминантные гены содержатся в генотипе собаки. О наличии некоторых рецессивных генов, которые не повлияли на окрас, но которые присутствуют в генотипе, мы можем судить по окрасам родителей этой собаки, а так же по тому, какие окрасы производит данная собака в вязке с теми или иными партнерами. То есть, только опытным путем. К примеру, у нас есть черная собака, полученная от шоколадного отца и красной матери. Мы можем с уверенностью говорить, что в генотипе данной собаки обязательно есть красный ген и шоколадный ген. Но если мы имеем черную собаку от двух черных родителей, информация о рецессивных генах, присутствующих в генотипе этой собаки, нам будет недоступна. Выяснить носительство в этом случае можно только в вязках с партнерами разных окрасов.
Знание генотипа племенных собак сильно облегчает жизнь заводчику, а так же предотвращает "артефакты" в разведении (например, когда от двух голубых собак "неожиданно" рождается черный щенок, что на деле говорит о том, что сука была повязана еще с кем-то).
Окрас шар-пея, как мы его видим, имеет свой генетический код, записанный в хромосоме. Хромосома – это носитель генетической информации. Она представляет собой уникальный для каждой собаки набор генов. Все гены в ней разбиты по парам. Эти пары называются "сериями". Серии в книгах по генетике принято обозначать буквами латинского алфавита: A , B , C , D , E , K (указаны только те серии, которые нужны для понимания окраса у шар-пея). Каждая пара (серия) отвечает за какой-то один признак, влияющий на формирование окраса собаки. А совокупность признаков дает результат в виде окраса, который мы видим у собаки. В каждой серии есть как минимум 2 гена, называемые "аллелями". Аллели – это своеобразные "мутации" одного исходного гена, произошедшие в ходе эволюции. В каждой серии есть доминантный ген (подавляющий проявление других генов своей серии) и рецессивный (подавляемый ген, который может проявиться только в том случае, если в паре с ним в хромосоме окажется точно такой же подавляемый ген). Гены влияют не только на стоящие с ними в одной паре, но и могут оказывать воздействие на другие пары генов, из других серий. Так, например, пара дильютных генов из серии B влияет на черный ген из серии K , и в результате перед нами оказывается шоколадная собака.
Окрас конкретной собаки – это результат работы целого ряда пар генов из разных серий.
Чтобы перейти к описанию каждого окраса, нужно кратко пояснить назначения шести главных серий генов, которые влияют на формирование окраса у шар-пея (поэтому указываются не все гены каждой серии, а только те, что могут быть у шар-пея не бракованного окраса).
А – серия , отвечающая за чистоту рыжего окраса.
Доминантный ген А y – отвечает за все вариации рыжих окрасов (красный, олень, изабелла, в т.ч. и дильютные – абрикосовый и т.д.).
Рецессивный ген aw – дает соболиный окрас. В этой же серии далее находятся гены чепрачного, черно-подпалого и рецессивного черного окрасов. Первые два встречаются у шар-пеев бракованных окрасов. Последний в шар-пеях не зарегистрирован вообще. Если проследить мутации генов серии А, то доминантой будет сплошной рыжий окрас, далее – зонарный соболиный, далее чепрачный, потом черно-подпалый, и, наконец, рецессивный сплошной черный.
В – серия , отвечающая за появление шоколадного пигмента взамен черному. Фактически, эта серия отвечает за все дильютные окрасы, имеющие шоколадную или лиловую маску, либо шоколадный или лиловый нос.
Ген В – доминантный ген, дающий черный пигмент. Черный нос, или черная маска на морде, черные кончики волос у соболиного окраса, черные зоны у черно-подпалых собак – это все работа доминантного гена B. Благодаря ему формируются все т.н. основные (не дильютные) окрасы, включая голубой и изабелловый.
Ген b – рецессивный ген, или так называемый "шоколадный ген", дающий шоколадный пигмент, взамен черного. Так как ген рецессивный, то проявление его возможно только в паре с таким же рецессивным геном. Наличие пары bb приводит к трансформации черного пигмента кожи и шерсти в шоколадный. Шоколадные маски, шоколадные носы, шоколадные кончики волос у соболиных дильютов, шоколадные зоны у шоколадно-подпалых собак – все это работа пары bb. От этой пары зависят такие окрасы как шоколадный, лиловый, абрикосовый дильют, красный дильют, кремовый дильют, изабелловый дильют.
С – серия , отвечающая за интенсивность окраса.
Доминантный ген C – это насыщенный окрас (например, ярко красный).
Рецессивный ген cch – осветленный окрас (кремовый, например).
D – серия , отвечающая за образование голубого, лилового, изабеллового и изабеллового дильютного окрасов.
Доминантный ген D – это ген черного пигмента (у "рыжих" собак). Он не влияет на красный пигмент.
Рецессивный ген d – так называемый "голубой ген", отвечающий за голубой окрас и его производные (изабелла, лиловый, изабелла дильют). Фактически, так же, как и "шоколадный ген", "голубой ген" влияет только на черный пигмент, трансформируя его в различные оттенки серого. Все, что в собаке обычного окраса будет черным, у собаки с двойным "голубым геном" (пара dd ) будет серым (голубым). Например, черная маска у оленьей собаки трансформируется в серую у шар-пея с парой голубых генов. При этом сама собака будет уже не оленьей, а изабелловой.
Е – серия , отвечающая за наличие распространения черного пигмента по телу собаки.
Доминантный ген Em – ген дающий маску на морде, темный язык, темные подушки лап, анус; причем, у собаки основного окраса это будет черная маска, синий язык, у собаки с двойным шоколадным геном – шоколадная маска, насыщенный лиловый язык, у собак с двойным голубым геном – голубая маска и синий язык.
Рецессивный ген E – это ген умеренного распространения черного пигмента по телу. У собаки может быть черный (шоколадный, голубой) нос, но при этом не будет маски. Когти могут быть светлые.
Самый хитрый ген из этой серии, и самый рецессивный ген e – он запрещает проявление черного пигмента вообще. Фактически, ген е "запрещает" проявление черного пигмента, даже если в генотипе есть доминантные гены черного пигмента и черного окраса (гены B , D , K ). Рецессивную пару ee могут иметь только светлые окрасы с полностью отсутствующей черной, шоколадной или голубой пигментацией. То есть, это полностью "депигментированные" окрасы. Несмотря на разницу в генотипах со светлыми окрасами, имеющими двойной шоколадный ген, они имеют те же названия, то есть, красный дильют, абрикосовый дильют, кремовый дильют, изабелловый дильют. Но при этом язык у собак с двойным ее остается довольно темным. Этот рецессивный ген часто играет с заводчиками дурную шутку, т.к. многие из них не понимают разницу между дильютными окрасами, полученными за счет шоколадных генов, и депегментированными окрасами, полученными за счет генов, запрещающих черный пигмент, поскольку и те, и другие в реестре окрасов называются "дильютными". Как результат – в вязке двух внешне дильютных собак можно получить черных и рыжих масковых щенков.
К – серия черного окраса.
Доминантный ген К ответственен за черный, голубой, лиловый, шоколадный окрасы. Но так же он может содержаться в генотипе собак, имеющих пару рецессивных генов ее. Так как эти гены просто блокируют проявление черного пигмента, доминирующий черный ген Кскрывается от наших глаз, и собака фенотипически может выглядеть кремовой дильютной, абрикосовой дильютной, и т.д.
Рецессивный ген k – это все обычные нечерные окрасы: красный, олень, изабелла и прочее.
Чтобы понять, как работают все эти гены вместе, рассмотрим генотип каждого окраса. Запись генотипа окраса делается в одну строку, гены указываются попарно, если мы знаем и те гены, которые дают фенотип (окрас), и те, о скрытом носительстве которых мы уже осведомлены. Но если мы точно знаем только фенотип собаки, и не в курсе относительно того, что эта собака несет в скрытом виде, мы пишем только доминантные гены, а вместо парного гена пишем прочерк, т.к. фактически на фенотип собаки этот рецессивный ген не влияет.
Итак:
Черный
Генотип: Ay - B - C - D - Em - K -, либо aw - B - C - D - Em - K -
(либо то же самое, но вместо Em - присутствует E - )
Поскольку черный окрас – это результат действия доминантных генов всех серий, он может оказаться носителем генов разнообразных окрасов. Поэтому получение от черного шар-пея пометов со щенками различных окрасов не должно вызывать удивления. Так, например, черная собака, имеющая такой генотип: AyawBbCcDdEmeKk с разными, правильно подобранными партнерами будет давать абсолютно все допустимые окрасы.
Как уже было сказано, на наличие в генотипе определенных рецессивных генов окраса (дильютного, голубого, красного) часто указывает отлив шерсти у собаки. Сейчас не редко можно видеть собак, чей окрас можно было бы классифицировать как очень темный "горький" шоколад с черной маской. Однако, это генетически черные собаки, и регистрировать их следует как черных, поскольку в вязках с шоколадными дильютными собаками они будут давать как шоколадных, так и черных щенков. Черные собаки, один из родителей которых – голубой или изабелла, могут давать голубоватый отлив по шерсти. Но опять-таки, это не повод записывать их как голубых.
Следует иметь в виду, что черный окрас не может быть получен от двух пигментированных красных, оленьих, изабелловых собак, поскольку у них отсутствует необходимый черный ген К. Точно так же черный не может быть получен от собак с двойным шоколадным геном или двойным голубым геном. То есть, к примеру, от двух шоколадных собак или от двух голубых собак не рождаются черные щенки.
Просмотров:
290
|
Добавил:
k-9
|
Дата:
05.04.2018
|
Шоколадный дильют
Генотип: Ay - bb C - D - Em - K -, либо aw - bb C - D - Em - K -
(либо то же самое, но вместо Em - присутствует E - )
Это "дильютная производная" от черного окраса. С одинаковым успехом его можно было бы назвать "черный дильют". Разница между шоколадным и черным окрасом – лишь на уровне одной пары генов из серии B. У черного окраса в этой серии всегда есть хотя бы один доминантный ген B , а у шоколадного дильютного шар-пея всегда только два рецессивных гена bb. Это означает, например, что от двух шоколадных собак не могут родиться черные щенки или другие щенки основных окрасов – красный, олень, крем, изабелла. Точно так же от двух шоколадных собак не могут родиться голубые щенки. Случаи, когда в помете от двух шоколадных собак рождается черный, красный, голубой, изабелловый щенки, по всей видимости, следует считать артефактом, и, очевидно, сука была довязана или сама довязалась с другим кобелем, что делает регистрацию помета в РКФ невозможной. В то же время, из-за наличия в генотипе доминантного гена черного окраса – К, - шоколадный окрас не может быть получен от двух пигментированных красных, оленьих, изабелловых собак, у которых этот ген отсутствует.
Голубой
Генотип: Ay - B - C - dd Em - K -, либо aw -B- C - dd Em - K -
(либо то же самое, но вместо Em - присутствует E - )
Голубой окрас отличается от черного только по паре генов в серии D. У черного шар-пея в этой паре присутствует доминантный ген D , который отвечает за полноценное формирование черного пигмента в шерсти. А у голубой собаки этого гена нет, но есть два рецессивных гена dd , которые в паре дают "размытый" черный пигмент, который человеческому глазу видится как голубой. По этой причине голубой окрас так же как и шоколадный называют "дильютным". Однако, природа возникновения голубого и шоколадного окраса различны. Поэтому при вязке голубого и шоколадного шар-пеев можно получить полностью черный помет.
Следует так же помнить, что отсутствие в генотипе голубых собак доминантного гена D делает невозможным рождение черных или иных окрасов, не входящих в группу "голубых" от двух голубых собак. То есть, два голубых, голубой и изабелловый, два изабелловых, голубой и лиловый, изабелловый и лиловый, два лиловых шар-пея на генетическом уровне не способны дать черных, красных, кремовых (с черным носом), шоколадных, оленьих, абрикосовых, крем-дильютных (без лилового пигмента), красных дильютных, соболиных или соболиных дильютных щенков. В вязке двух голубых собак могут рождаться только: голубые, изабеллы, лиловые и изабелловые дильюты. Так же в вязке этих собак могут рождаться кремовые щенки с голубой или лиловой пигментацией, но в виду отсутствия специальных названий для этих окрасов, их так же регистрируют как крем и крем дильют.
Как и в случае с шоколадным окрасом, из-за наличия в генотипе доминантного гена черного окраса – К, - голубой окрас не может быть получен от двух пигментированных красных, оленьих, изабелловых собак.
Лиловый
Генотип: Ay - bb C - dd Em - K - или aw - bb C - dd Em - K -
(либо то же самое, но вместо Em - присутствует E - )
Лиловый окрас получается при комбинации двух пар рецессивных генов – bb и dd. Каждая из пар "отвечает" за свой окрас. Первая – за шоколадный. Вторая – за голубой. Но когда эти пары оказываются вместе в одной хромосоме, возникает лиловый окрас. Формально, лиловую собаку можно назвать одновременно шоколадной и голубой. От голубого она отличается только тем, что к имеющимся голубым генам добавляются и шоколадные гены. А от шоколадной собаки тем, что к имеющимся шоколадным генам добавляется пара голубых генов.
Обратите внимание на то, что от черного окраса лиловый отличается по генотипу только наличием рецессивных генов в B и D сериях. В остальном – все то же самое. То есть, лиловый – это окрас, несущий доминантный черный ген K. Точно так же, как носителями черного гена являются черный, шоколадный и голубой окрасы. Без этого черного гена возникновение этих четырех окрасов (черного, голубого, шоколада и лилового) было бы невозможным. Из этого вытекает следующее правило: рыжие собаки с черным пигментом (они не имеют черного гена K в своем генотипе) в вязке друг с другом не способны родить лиловых, шоколадных, голубых или черных щенков. При появлении таких щенков в помете у рыжих масковых собак, следует поставить под сомнение происхождение этих щенков (как минимум – поставить под сомнение отцовство). Следует так же помнить, что, поскольку лиловые собаки не несут в своем генотипе доминантных генов B , то в вязке двух лиловых собак, невозможно получить голубых собак, т.к. генотип голубой собаки требует, чтобы собака имела доминантный ген B , а у лиловых собак его просто нет. Две лиловых собаки могут дать только лиловых щенков или изабелловый дильют, на большее они не способны в силу своего генотипа. Вязка лилового шар-пея с шоколадным так же не может дать черных, красных, оленьих или голубых щенков, поскольку оба окраса лишены доминантного гена B , необходимого для формирования основных окрасов. То есть, при вязке шоколада, не несущего голубой ген, с лиловым получается полностью дильютный помет (но не голубые!). В самом простом случае все щенки будут шоколадными. Лиловый и голубой, лиловый и изабелловый в паре всегда будут давать как минимум голубых щенков, а при наличии в генотипе партнера скрытых шоколадного (у голубой и изабеллы) и красного (у голубой) генов – еще и лиловый, изабеллу и изабелловый дильют. Вязка лилового и изабеллового дильюта не даст ничего кроме лиловых и изабелловых дильютов (второй окрас – в том только случае, если лиловый шар-пей несет в себе рецессивный ген k – ген, "не черного" окраса).
Красный
Генотип красной собаки с маской: Ay - B - C - D - Em - kk
Генотип красной собаки без маски, но с черным носом: Ay - B - C - D - E - kk
Красный окрас так же относится к основным (не дильютным окрасам). Отличие от черного окраса на генетическом уровне – в серии генов K. У красной собаки с черным пигментом ген черного окраса шерсти отсутствует! Это означает, что две красные масковые собаки не могут произвести на свет окрасы, образование которых требует наличия в генотипе доминантного гена K : черный, голубой, шоколадный, лиловый. При рождении шоколадного, черного, голубого, лилового щенка от двух красных родителей (с маской или с черными носами) следует поставить под сомнение происхождение помета, в котором они родились. Заводчику следует рекомендовать проведение генетической экспертизы для установления отцовства.
Красный дильют
На генетическом уровне красный дильютный окрас может быть получен двумя совершенно разными способами.
Вариант 1:
Генотип красного дильюта с шоколадной маской: Ay - bb C - D - Em - kk
Генотип красного дильюта с шоколадным носом (без маски): Ay - bb C - E - kk
Генотип красного дильюта без шоколадного пигмента: Ay - bb C - D - eekk или Ay - bb C - D - eeK -
Отличие этого варианта красного дильютного окраса от просто красного такое же, как отличие шоколадного окраса от черного или лилового от голубого – красный дильютный окрас не имеет доминантного гена B в своем генотипе, а его место занимает рецессивная "шоколадная" пара bb. У красного дильютного шар-пея, полученного в таком сочетании генов, может быть шоколадный нос или даже шоколадная маска на морде, шоколадный анус и подушечки лап. Отсутствие шоколадного пигмента на теле тоже возможно, за счет работы еще одной пары рецессивных генов – ee , запрещающих распространение темного пигмента по телу. При этом язык у такой собаки будет светлым лавандовым. В вязке с шоколадным окрасом такая собака никогда не даст черных или других основных окрасов, а так же не даст голубых. В вязке с лиловым партнером такая красная дильютная собака даст только дильютных щенков, в самом простом случае – шоколадных. Для удобства этот вариант окраса, полученный под действием генов bb называют "истинным дильютом".
Вариант 2:
Генотип красного дильюта без шоколадного пигмента: Ay - B - C - D - eeK - или Ay - B - C - D - eekk ,
либо aw - B - C - D - eeK - или aw - B - C - D - eekk
Внешне эта красная дильютная собака выглядит совершенно однотонной, без малейшего намека на шоколадный налет на морде, подушках, анусе. При этом у нее довольно темный лавандовый язык, что часто принимается за очень хорошую пигментацию. Обратите внимание, что собаки с генотипом Ay - B - C - D - eeK - и aw - B - C - D - eeK - имеют черный ген! Это означает, что они могут давать черных щенков и другие окрасы, образование которых требует наличия этого гена. К примеру, с красной масковой собакой такая собака может дать черных щенков (а так же шоколад, голубых или лиловых, если в генотипе обоих есть соответствующие рецессивные гены). А с шоколадным партнером может так же дать полностью черный помет.
Красная дильютная собака с генотипом Ay - B - C - D - eekk или aw - B - C - D - eekk не будет давать черных (шоколад и прочее) щенков в вязке с красной (оленьей, изабелловой) масковой собакой, т.к. у нее нет черного гена K. При вязке с шоколадным или лиловым партнером от такой собаки можно получить полностью черный помет. Поэтому красный дильютный окрас, образованный таким генотипом, часто называют "псевдодильют", тем самым указывая на то, что этот внешне 100% дильютный окрас не имеет в генотипе пары шоколадных (дильютных) генов bb. В лучшем случае такая собака является носителем рецессивного скрытого гена b. В худшем – не несет и никогда не даст шоколадных щенков.
Следует так же обратить внимание, что при наличии пары генов ee становится невидимой и пара генов соболиного окраса, т.к. пара ее маскирует проявление черного пигмента в волосах (т.е. черные кончики волосков осветляются до светлого рыжего).
Из-за сложности в определении генотипа красной дильютной собаки, единственным способом проверки его является опыт. То есть, вязка с шоколадным партнером укажет на то, имеет ли красная дильютная собака рецессивный ген b и сколько – один или два (если генов два – помет будет полностью дильютный, если один – помимо шоколадных щенков могут родиться щенки основных окрасов, а если ген отсутствует – все основные окрасы). А вязка с красным масковым партнером укажет на наличие или отсутствие гена K , формирующего черный окрас (этот ген подавляется парой ee , поэтому его "не видно"). Если ген отсутствует – весь помет будет рыжий. Если присутствует – часть или весь помет будет черный.
Олений
Генотип: Ay - B - CcchD - Em - kk
(либо то же самое, но вместо Em - присутствует E - )
По всей видимости, на образование этого окраса влияет серия генов C. Ген насыщенного окраса C не полностью доминирует над геном осветления окраса cch , поэтому если они находятся в паре, окрас из насыщенного красного превращается в "блеклый" олений. Так же существует теория о существовании генов модификаторов, которые влияют на насыщение красного окраса. В поддержку этой теории говорит и то обстоятельство, что от двух оленьих собак невозможно получить махагоновых щенков, даже если обе оленьи собаки имеют по одному махагоновому родителю. В то время как махагоновая собака в вязке с практически любой рыжей собакой может дать махагоновых щенков.
На генетическом уровне олений окрас отличается от черного окраса тем, что имеет рецессивную пару kk , то есть, не имеет черного гена. Вязка двух оленьих шар-пеев исключает возможность рождения черных, шоколадных, голубых, лиловых щенков, т.к. для получения этих окрасов нужен доминантный черный ген К, который у оленьих собак отсутствует. В вязке двух оленьих собак, имеющих голубой ген, могут родиться изабелловые щенки. В вязке оленьих собак, имеющих дильютный (шоколадный) ген, могут родиться абрикосовые дильютные щенки.
Абрикосовый дильют
По аналогии с красным дильютным окрасом, точно так же существует два варианта возникновения этого окраса. То есть, так же, как и красный дильют, абрикосовый дильют может быть "истинным" и "псевдо". И соответственно либо иметь рецессивную пару шоколадных генов bb , либо вообще не иметь шоколадного гена (не иметь возможности давать шоколадных щенков), и быть носителем черного гена К.
Вариант абрикосового дильюта с шоколадным пигментом:
Генотип абрикосового дильюта с шоколадной маской: Ay - bb CcchD - Em - kk
Генотип красного дильюта с шоколадным носом (без маски): Ay - bb CcchE - kk
Генотип красного дильюта без шоколадного пигмента: Ay - bb CcchD - eekk или Ay - bb CcchD - eeK -
Вариант абрикосового дильюта депегментированного:
Ay - B - CcchD - eeK - и Ay - B - CcchD - eekk ,
либо aw - B - CcchD - eeK - и aw - B - CcchD - eekk
Для подробностей смотрите описание красного дильютного окраса. Все те же ограничения и те же способы проверки генотипа. То есть, к примеру, вязка депегментированного абрикосового дильюта с красным масковым или с шоколадным партнером может дать черный помет. Вязка абрикосового дильюта с шоколадным пигментом не даст черных щенков с шоколадным партнером, и т.п., описанное для красного дильютного окраса.
Просмотров:
340
|
Добавил:
k-9
|
Дата:
05.04.2018
|
Кремовый
Генотип: Ay - cchcchB - D - Em - kk
(либо то же самое, но вместо Em - присутствует E - )
Отличие от красного и оленьего окрасов – в наличии пары рецессивных генов cchcch , которые ослабляют красный пигмент до очень светлого.
От двух кремовых собак невозможно получить ярких красных или оленьих. Крем с кремом дает только крем (или крем дильют).
Кремовый дильют
Вариант кремового дильюта с шоколадным пигментом:
Генотип: Ay - cchcchbbD - Em - kk или Ay - cchcchbbD - EEkk
Вариант кремового дильюта депигментированного:
Генотип: Ay - cchcchB - D - eekk или aw - cchcchB - D - eekk
либо Ay - cchcchB - D - ee К- или aw - cchcchB - D - ee К-
Таким образом, так же как и абрикосовый дильют, и красный дильют, этот окрас может быть истинным дильютным (полученным за счет шоколадных генов bb ), и псевдо дильютом, полученным за счет пары генов ee. Генотипическое отличие крем-дильюта от крема будет точно таким же, как оленя от абрикосового дильюта и красного от красного дильюта. Ограничения и проверка генотипа – такие же.
В случае получения кремового дильюта за счет генов bb , на морде видна легкая шоколадная маска или напыление. В случае если кремовый дильют получен за счет генов ee – это почти белая собака, без тени намека на шоколадный пигмент, но при этом имеющая более темный, чем можно ожидать, язык.
Если кремовый дильютный шар-пей рожден от двух шоколадных собак, то в его генотипе всегда будет два гена bb , но при этом он может содержать ген черного окраса К и давать черных щенков в комбинации с рыжими собаками.
Изабелловый
Генотип:
Насыщенная ("красноватая") изабелла с серой маской: Ay - B - C - dd Em - kk
Насыщенная ("красноватая") изабелла без маски, с серым носом: Ay - B - C - dd E - kk
Светлая ("оленья") изабелла с серой маской: Ay - B - Ccchdd Em - kk
Светлая ("оленья") изабелла без маски, с серым носом: Ay - B - Ccchdd Em - kk
"Соболиная" изабелла с серой маской: awaw B - C - dd Em - kk
"Соболиная" изабелла без маски, с серым носом: awaw B - C - dd Em - kk
На данном этапе нет смысла выделять соболиную изабеллу в отдельный окрас, т.к. явление это редкое. Однако следует учитывать возможность появления в пометах и таких щенков.
На генетическом уровне изабелловый окрас можно описать как одновременно голубой и рыжий (красный или олений). Отличие от голубого окраса точно такое же, как отличие красного или оленьего от черного – у изабеллы отсутствует ген черного окраса К.
Следует помнить, что изабелловый окрас может быть получен ТОЛЬКО от ДВУХ голубогенных собак. При этом одна из собак должна быть не-дильютной (т.е. не иметь пары шоколадных генов), т.к. для формирования изабеллы нужен доминантный ген B , как и для любого основного "рыжего" окраса. Если в родословных производителей нет голубых (изабелловых, лиловых) собак, и если производители не проверены на голубых собаках (не давали голубых щенков), не следует регистрировать щенков от таких родителей как изабеллу. В том случае, если заводчик настаивает на том, что в помете рожден именно изабелловый щенок – необходимо предоставить описание экспертом этого щенка после первой линьки, когда окрас уже очевиден, и только после этого можно вносить поправку к окрасу данной собаки в общепометную карту и в родословную.
Изабелловый дильют
Вариант изабеллового дильюта с лиловым пигментом:
Генотип изабеллового дильюта с лиловой маской: Ay - bb C - dd Em - kk
Генотип изабеллового дильюта с лиловым носом: Ay - bb C - dd E - kk
Вариант изабеллового дильюта без пигмента:
Генотип изабеллового дильюта депигментированного (нет лилового пигмента): Ay - B - C - dd ее kk
либо Ay - B - C - dd ееК-
В первом варианте на генетическом уровне изабелловый дильют отличается от изабеллы тем, что имеет пару рецессивных шоколадных генов bb. Эти гены и трансформируют весь серый (ослабленный голубыми генами dd ) пигмент в лиловый. Получить этот изабелловый дильют можно ТОЛЬКО от собак, в генотипе которых точно есть рецессивные гены голубого, шоколадного и красного (не-черного) окрасов. То есть, оба производителя должны иметь как минимум по одному гену: b , d , k.
Во втором варианте разница между изабеллой и изабелловым дильютом в рецессивной паре ee , которая делает проявление серого пигмента невозможным. Встречается такая разновидность окраса крайне редко, поскольку и сам окрас пока еще чрезвычайно редок.
Соболиный
Генотип соболиного окраса определяется парой генов aw , рецессивных по отношению к сплошному рыжему:
Яркий ("красный") соболь с маской: awawB - C - D - Em - kk
Яркий ("красный") соболь без маски, но с черным носом: awawB - C - D - E - kk
Светлый ("олений") соболь с маской: awawB - CcchD - Em - kk
Светлый ("олений") соболь без маски, но с черным носом: awawB - CcchD - E - kk
Соболиный дильют
Яркий ("красный") соболь с шоколадной маской и шоколадной зонарностью: awawbbC - D - Em - kk
Яркий ("красный") соболь без маски, но с шоколадным носом и шоколадной зонарностью: awawbbC - D - EEkk
Светлый ("олений") соболь с шоколадной маской: awawbbCcchD - Em - kk
Светлый ("олений") соболь без маски, но с шоколадным носом: awawbbCcchD - EEkk
Несмотря на то, что официально этот окрас еще не регистрируют, знание генотипа окраса поможет заводчикам правильно его идентифицировать и применять в разведении. Отличие соболиного дильюта от соболя состоит лишь в наличии пары генов bb. "Псевдо дильютных" соболей не существует по той причине, что пара генов ee скрывает любое вообще проявление темного пигмента, поэтому присутствие этой пары генов в генотипе собаки делает ее совершенно идентичной обычному красному дильюту или абрикосовому дильюту.
Внешне соболиный дильют выглядит более ровным, нежели основной соболиный, поскольку весь черный пигмент в нем трансформируется в шоколадный, и как результат – контраст между рыжей и шоколадной зонами в остевых волосах гораздо меньше заметен. В то же время, при сравнении красной дильютной или абрикосовой дильютной собак с соболиной дильютной разница в окрасе становится совершенно очевидна. Соболиные дильютные окрасы могут рождаться практически шоколадными, и "перецветать" по мере роста и смены шерсти. При рассматривании остевого волоса легко заметить переходы от светлого к шоколадному окрасу. Спина может иметь более или менее выраженный шоколадный налет. В вязках с шоколадными собаками этот окрас ведет себя так же, как и любой другой дильютный окрас (истинный).
IV. От генотипа к фенотипу. Артефакты в разведении.
Несмотря на то, что генетика – это вероятностная наука, и не все в ней можно просчитать заранее, тем не менее, существуют некоторые аксиомы в разведении окрасов, не подвергающиеся сомнению. Проще говоря, есть правила, которые не нарушаются никогда.
Правило 1: от двух черных собак можно получить любые окрасы, если они заложены в генотипе в виде рецессивных генов.
Правило 2: от двух красных (оленьих) собак с масками (с черными носами) нельзя получить шоколадных, черных, голубых, лиловых щенков, поскольку у красных (оленьих) собак нет в генотипе доминантного черного гена К, без которого черный, шоколадный, голубой и лиловый окрасы невозможны.
Правило 3: от двух шоколадных собак нельзя получить черных, голубых, красных (оленьих), кремовых или изабелловых щенков, т.к. у шоколадных собак в генотипе отсутствует ген черного пигмента В, отвечающий за наличие этого пигмента в фенотипе собаки (маска или черный нос у красных, полностью черный волос у черных, серый волос у голубых).
Правило 4: от двух голубых, от голубой и изабеллы, голубой и лилового собак получаются ТОЛЬКО окрасы с двойным голубым геном, т.е. голубые, изабелла, лиловые. Получение черных, шоколадных, красных и оленьих с маской или черным носом щенков невозможно по причине отсутствия доминантного гена D у голубых, изабеллы и лиловых собак.
Правило 5: от двух собак с шоколадным пигментом невозможно получение собак с черным или серым (голубым) пигментом.
Просмотров:
319
|
Добавил:
k-9
|
Дата:
05.04.2018
|
Правило 6: от двух кремовых собак можно получить только кремовых щенков (или кремовых дильютных). От двух кремовых дильютных собак можно получить ТОЛЬКО кремовый дильют.
Правило 7: от двух изабелловых собак нельзя получить голубых или лиловых щенков. Возможно получение только изабеллы и изабеллового дильюта.
Правило 8: от двух лиловых собак нельзя получить голубых, изабелловых или шоколадных щенков. Можно получить только лиловых и изабелловый дильют.
Правило 9: от двух изабелловых дильютных собак можно получить ТОЛЬКО изабелловый дильют.
Правило 10: от двух соболиных собак нельзя получить щенков сплошного оленьего или красного окраса, т.к. эти окрасы доминантны по отношению к соболю.
Правило 11: от шоколадной и лиловой собаки нельзя получить черный или любой иной окрас с черным или серым пигментом.
Правило 12: от собак с серым (голубым) ли лиловым пигментом возможно получение кремовых щенков с серым (голубым) или лиловым пигментом, которые на данном этапе регистрируются как крем или кремовый дильют в виду отсутствия собственных названий.
Это лишь несколько основных правил, которые позволяют избежать ошибок в разведении. При получении в пометах "артефактов" рекомендуется проведение независимой генетической экспертизы для установления отцовства и материнства, либо помет не подлежит регистрации в виду сомнительности его происхождения.
Просмотров:
262
|
Добавил:
k-9
|
Дата:
05.04.2018
| |
|
|
| Вход на сайт |
|
|
| Поиск |
|
|
| Календарь |
| « Апрель 2018 » |
| Пн |
Вт |
Ср |
Чт |
Пт |
Сб |
Вс |
| | | | | | | 1 | | 2 | 3 | 4 | 5 | 6 | 7 | 8 | | 9 | 10 | 11 | 12 | 13 | 14 | 15 | | 16 | 17 | 18 | 19 | 20 | 21 | 22 | | 23 | 24 | 25 | 26 | 27 | 28 | 29 | | 30 | |
|
| Архив записей |
|
|
| Друзья сайта |
|
|
|




